

Apela promotes blood vessel regeneration and remodeling in zebrafish
- PMID: 38355946
- PMCID: PMC10867005
- DOI: 10.1038/s41598-023-50677-1
Apela promotes blood vessel regeneration and remodeling in zebrafish
Abstract
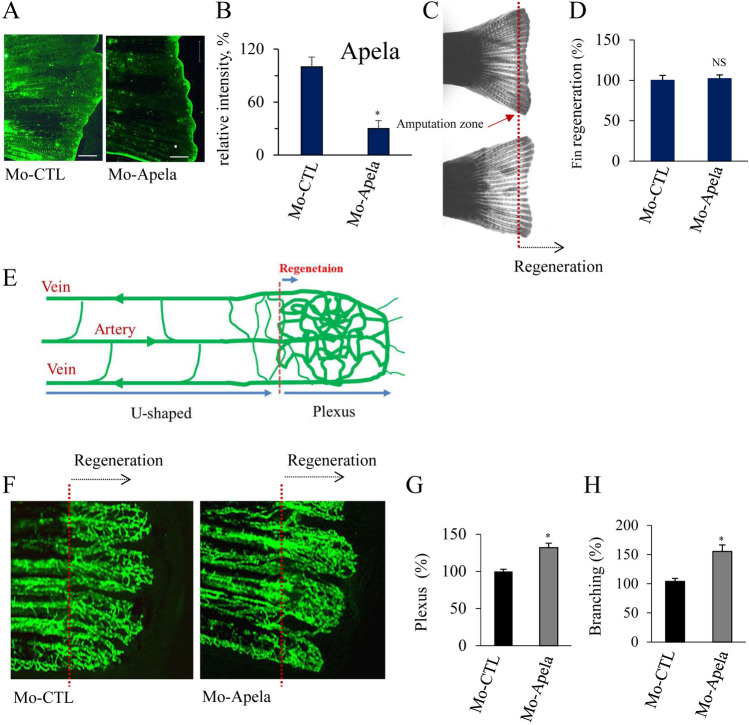
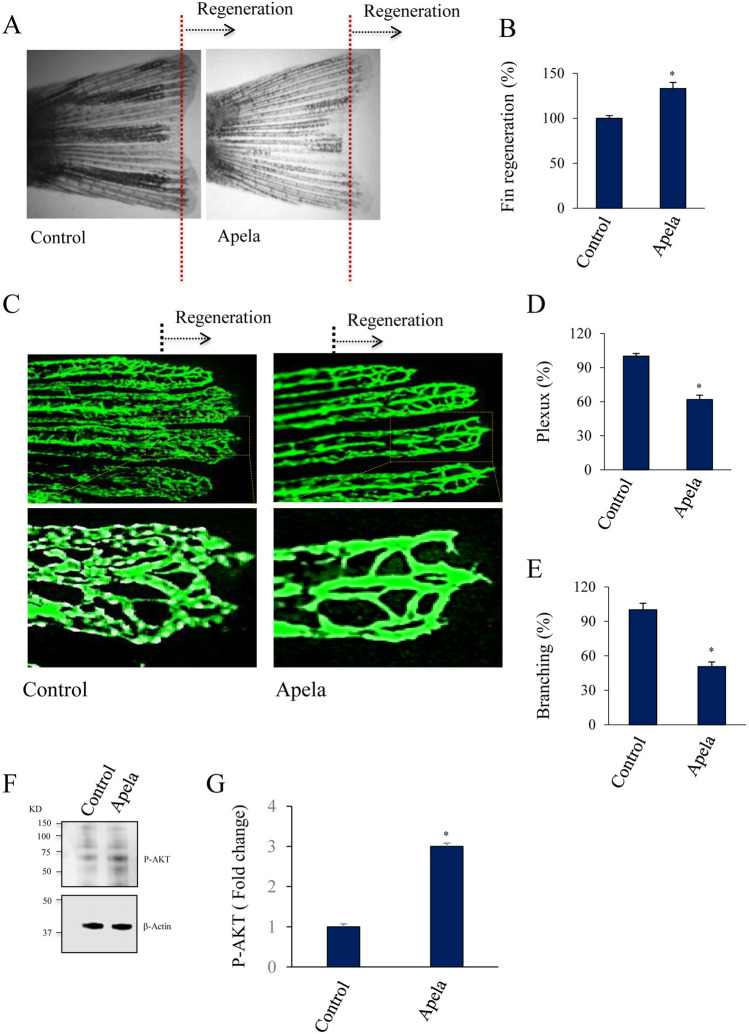
In contrast to adult mammals, zebrafish display a high capacity to heal injuries and repair damage to various organs. One of the earliest responses to injury in adult zebrafish is revascularization, followed by tissue morphogenesis. Tissue vascularization entails the formation of a blood vessel plexus that remodels into arteries and veins. The mechanisms that coordinate these processes during vessel regeneration are poorly understood. Hence, investigating and identifying the factors that promote revascularization and vessel remodeling have great therapeutic potential. Here, we revealed that fin vessel remodeling critically depends on Apela peptide. We found that Apela selectively accumulated in newly formed zebrafish fin tissue and vessels. The temporal expression of Apela, Apln, and their receptor Aplnr is different during the regenerative process. While morpholino-mediated knockdown of Apela (Mo-Apela) prevented vessel remodeling, exogenous Apela peptide mediated plexus repression and the development of arteries in regenerated fins. In contrast, Apela enhanced subintestinal venous plexus formation (SIVP). The use of sunitinib completely inhibited vascular plexus formation in zebrafish, which was not prevented by exogenous application. Furthermore, Apela regulates the expression of vessel remolding-related genes including VWF, IGFPB3, ESM1, VEGFR2, Apln, and Aplnr, thereby linking Apela to the vascular plexus factor network as generated by the STRING online database. Together, our findings reveal a new role for Apela in vessel regeneration and remodeling in fin zebrafish and provide a framework for further understanding the cellular and molecular mechanisms involved in vessel regeneration.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
Aplnra/b Sequentially Regulate Organ Left-Right Patterning via Distinct Mechanisms.Int J Biol Sci. 2019 May 11;15(6):1225-1239. doi: 10.7150/ijbs.30100. eCollection 2019. Int J Biol Sci. 2019. PMID: 31223282 Free PMC article.
-
Bone morphogenetic protein signaling promotes morphogenesis of blood vessels, wound epidermis, and actinotrichia during fin regeneration in zebrafish.FASEB J. 2015 Oct;29(10):4299-312. doi: 10.1096/fj.15-272955. Epub 2015 Jul 6. FASEB J. 2015. PMID: 26148971
-
Loss of Apela Peptide in Mice Causes Low Penetrance Embryonic Lethality and Defects in Early Mesodermal Derivatives.Cell Rep. 2017 Aug 29;20(9):2116-2130. doi: 10.1016/j.celrep.2017.08.014. Cell Rep. 2017. PMID: 28854362 Free PMC article.
-
Signaling networks organizing regenerative growth of the zebrafish fin.Trends Genet. 2015 Jun;31(6):336-43. doi: 10.1016/j.tig.2015.03.012. Epub 2015 Apr 27. Trends Genet. 2015. PMID: 25929514 Review.
-
The role of blood flow in vessel remodeling and its regulatory mechanism during developmental angiogenesis.Cell Mol Life Sci. 2023 May 23;80(6):162. doi: 10.1007/s00018-023-04801-z. Cell Mol Life Sci. 2023. PMID: 37221410 Free PMC article. Review.
Cited by
-
Repression of apelin Furin cleavage sites provides antimetastatic strategy in colorectal cancer.EMBO Mol Med. 2025 Mar;17(3):504-534. doi: 10.1038/s44321-025-00196-5. Epub 2025 Feb 17. EMBO Mol Med. 2025. PMID: 39962271 Free PMC article.
-
Neuronal activation in the axolotl brain promotes tail regeneration.NPJ Regen Med. 2025 May 8;10(1):22. doi: 10.1038/s41536-025-00413-2. NPJ Regen Med. 2025. PMID: 40341072 Free PMC article.
References
-
- Wacker A, Gerhardt H. Endothelial development taking shape. Curr. Opin. Cell Biol. 2011;23:676–685. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
