

The role of filamentous matrix molecules in shaping the architecture and emergent properties of bacterial biofilms
- PMID: 38358118
- PMCID: PMC10903470
- DOI: 10.1042/BCJ20210301
The role of filamentous matrix molecules in shaping the architecture and emergent properties of bacterial biofilms
Abstract
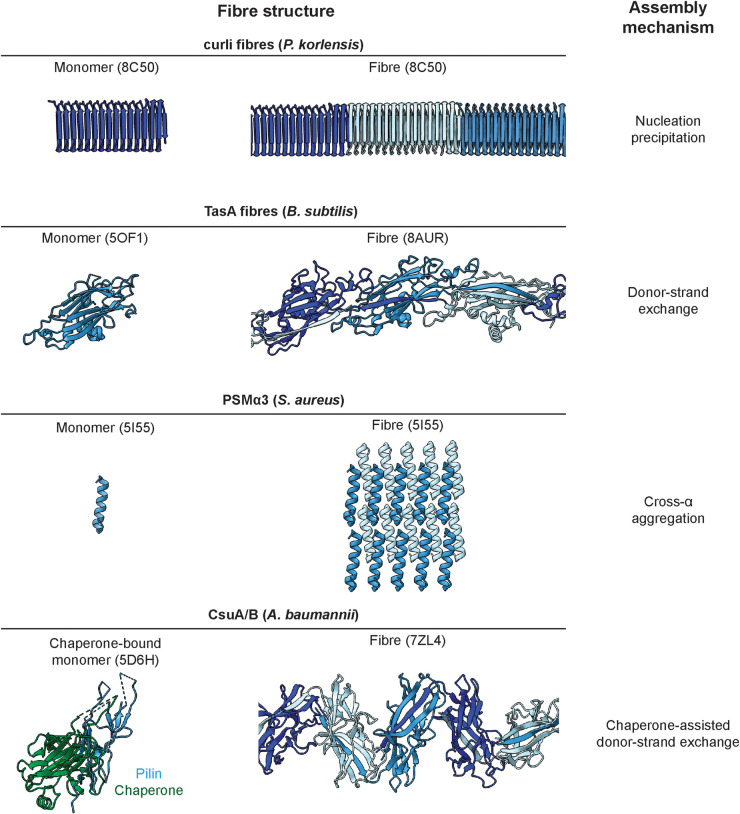
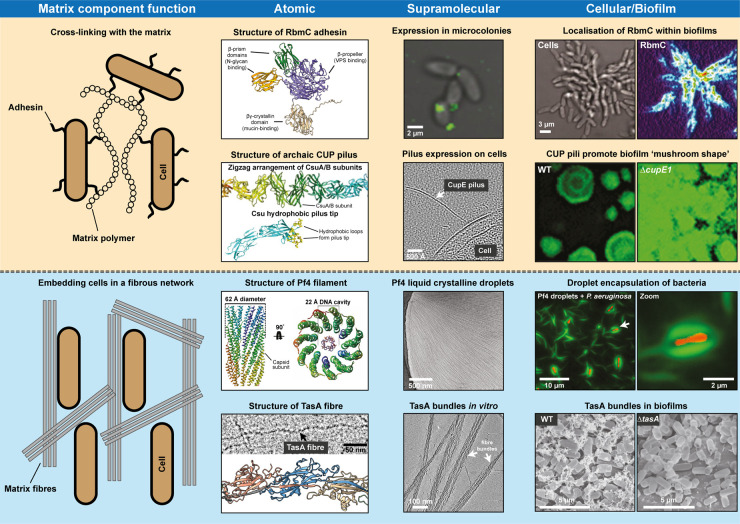
Numerous bacteria naturally occur within spatially organised, multicellular communities called biofilms. Moreover, most bacterial infections proceed with biofilm formation, posing major challenges to human health. Within biofilms, bacterial cells are embedded in a primarily self-produced extracellular matrix, which is a defining feature of all biofilms. The biofilm matrix is a complex, viscous mixture primarily composed of polymeric substances such as polysaccharides, filamentous protein fibres, and extracellular DNA. The structured arrangement of the matrix bestows bacteria with beneficial emergent properties that are not displayed by planktonic cells, conferring protection against physical and chemical stresses, including antibiotic treatment. However, a lack of multi-scale information at the molecular level has prevented a better understanding of this matrix and its properties. Here, we review recent progress on the molecular characterisation of filamentous biofilm matrix components and their three-dimensional spatial organisation within biofilms.
Keywords: bacteria; biofilms; extracellular matrix; microbiology; structural biology.
© 2024 The Author(s).
Conflict of interest statement
The authors declare that there are no competing interests associated with the manuscript.
Figures
Similar articles
-
Emergent Biological Endurance Depends on Extracellular Matrix Composition of Three-Dimensionally Printed Escherichia coli Biofilms.ACS Synth Biol. 2021 Nov 19;10(11):2997-3008. doi: 10.1021/acssynbio.1c00290. Epub 2021 Oct 15. ACS Synth Biol. 2021. PMID: 34652130 Free PMC article.
-
Biofilms: an emergent form of bacterial life.Nat Rev Microbiol. 2016 Aug 11;14(9):563-75. doi: 10.1038/nrmicro.2016.94. Nat Rev Microbiol. 2016. PMID: 27510863 Review.
-
Dynamic remodeling of microbial biofilms by functionally distinct exopolysaccharides.mBio. 2014 Aug 5;5(4):e01536-14. doi: 10.1128/mBio.01536-14. mBio. 2014. PMID: 25096883 Free PMC article.
-
Escherichia coli biofilms have an organized and complex extracellular matrix structure.mBio. 2013 Sep 10;4(5):e00645-13. doi: 10.1128/mBio.00645-13. mBio. 2013. PMID: 24023384 Free PMC article.
-
Biofilm Matrixome: Extracellular Components in Structured Microbial Communities.Trends Microbiol. 2020 Aug;28(8):668-681. doi: 10.1016/j.tim.2020.03.016. Epub 2020 Apr 21. Trends Microbiol. 2020. PMID: 32663461 Review.
Cited by
-
Biofilm-mediated bioremediation of xenobiotics and heavy metals: a comprehensive review of microbial ecology, molecular mechanisms, and emerging biotechnological applications.3 Biotech. 2025 Apr;15(4):78. doi: 10.1007/s13205-025-04252-2. Epub 2025 Mar 7. 3 Biotech. 2025. PMID: 40060289 Review.
-
Mapping the ultrastructural topology of the corynebacterial cell surface.PLoS Biol. 2025 Apr 15;23(4):e3003130. doi: 10.1371/journal.pbio.3003130. eCollection 2025 Apr. PLoS Biol. 2025. PMID: 40233127 Free PMC article.
-
Biofilm-Associated Amyloid Proteins Linked with the Progression of Neurodegenerative Diseases.Int J Mol Sci. 2025 Mar 17;26(6):2695. doi: 10.3390/ijms26062695. Int J Mol Sci. 2025. PMID: 40141340 Free PMC article. Review.
-
Architectural dissection of adhesive bacterial cell surface appendages from a "molecular machines" viewpoint.J Bacteriol. 2024 Dec 19;206(12):e0029024. doi: 10.1128/jb.00290-24. Epub 2024 Nov 5. J Bacteriol. 2024. PMID: 39499080 Free PMC article. Review.
-
Targeted disruption of phage liquid crystalline droplets abolishes antibiotic tolerance of bacterial biofilms.bioRxiv [Preprint]. 2025 Jul 28:2025.07.28.667140. doi: 10.1101/2025.07.28.667140. bioRxiv. 2025. PMID: 40766552 Free PMC article. Preprint.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources