

BRCA1 protein dose-dependent risk for embryonic oxidative DNA damage, embryopathies and neurodevelopmental disorders with and without ethanol exposure
- PMID: 38359745
- PMCID: PMC10877410
- DOI: 10.1016/j.redox.2024.103070
BRCA1 protein dose-dependent risk for embryonic oxidative DNA damage, embryopathies and neurodevelopmental disorders with and without ethanol exposure
Abstract
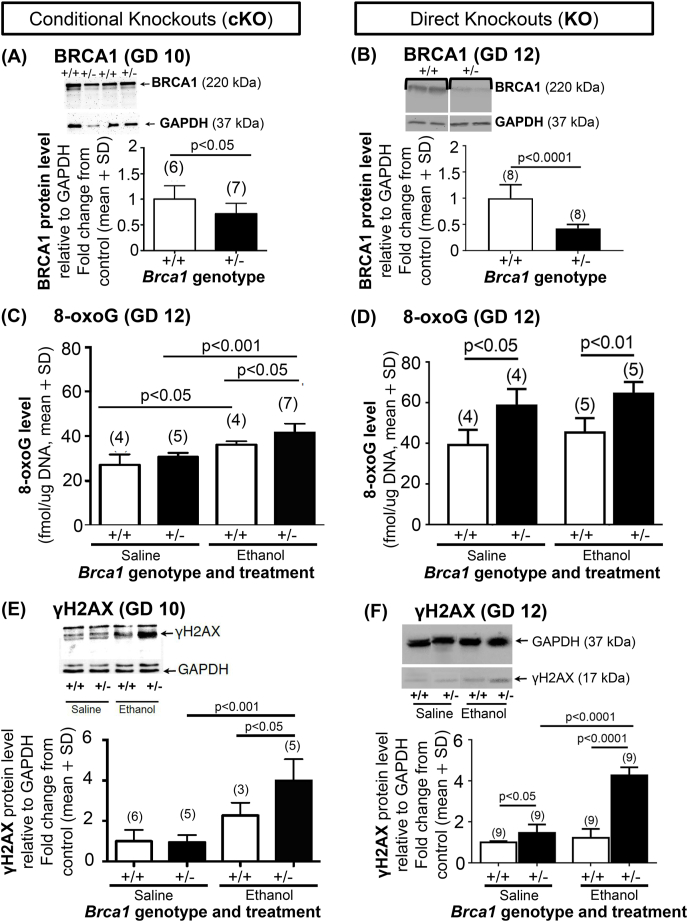
Although widely known as a tumor suppressor, the breast cancer 1 susceptibility protein (BRCA1) is also important in development, where it regulates fetal DNA repair pathways that protect against DNA damage caused by physiological and drug-enhanced levels of reactive oxygen species (ROS). We previously showed that conditional heterozygous (+/-) knockout (cKO) mouse embryos with a minor 28% BRCA1 deficiency developed normally in culture, but when exposed to the ROS-initiating drug, alcohol (ethanol, EtOH), exhibited embryopathies not evident in wild-type (+/+) littermates. Herein, we characterized a directBrca1 +/- knockout (KO) model with a 2-fold greater (58%) reduction in BRCA1 protein vs. the cKO model. We also characterized and compared learning & memory deficits in both the cKO and KO models. Even saline-exposed Brca1 +/- vs. +/+ KO progeny exhibited enhanced oxidative DNA damage and embryopathies in embryo culture and learning & memory deficits in females in vivo, which were not observed in the cKO model, revealing the potential pathogenicity of physiological ROS levels. The embryopathic EtOH concentration for cultured direct KO embryos was half that for cKO embryos, and EtOH affected Brca1 +/+ embryos only in the direct KO model. The spectrum and severity of EtOH embryopathies in culture were greater in both Brca1 +/- vs. +/+ embryos, and direct KO vs. cKO +/- embryos. Motor coordination deficits were evident in both male and female Brca1 +/- KO progeny exposed in utero to EtOH. The results in our direct KO model with a greater BRCA1 deficiency vs. cKO mice provide the first evidence for BRCA1 protein dose-dependent susceptibility to developmental disorders caused by physiological and drug-enhanced oxidative stress.
Keywords: Alcohol (ethanol, EtOH); Breast cancer 1 susceptibility gene (Brca1); Conditional knockout (cKO) mouse model; DNA damage and repair; Developmental disorders; Fetal alcohol spectrum disorders (FASD); Knockout (KO) mouse model; Reactive oxygen species (ROS).
Copyright © 2024 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest None.
Figures





References
-
- Daum H., Peretz T., Laufer N. BRCA mutations and reproduction. Fertil. Steril. 2018;109(1):33–38. - PubMed
-
- Lonning P.E., Berge E.O., Bjornslett M., Minsaas L., Chrisanthar R., Hoberg-Vetti H., Dulary C., Busato F., Bjorneklett S., Eriksen C., Kopperud R., Axcrona U., Davidson B., Bjorge L., Evans G., Howell A., Salvesen H.B., Janszky I., Hveem K., Romundstad P.R., Vatten L.J., Tost J., Dorum A., Knappskog S. White blood cell BRCA1 promoter methylation status and ovarian cancer risk. Ann. Intern. Med. 2018;168(5):326–334. - PubMed
-
- Wang Y., Bernhardy A.J., Cruz C., Krais J.J., Nacson J., Nicolas E., Peri S., van der Gulden H., van der Heijden I., O'Brien S.W., Zhang Y., Harrell M.I., Johnson S.F., Candido Dos Reis F.J., Pharoah P.D., Karlan B., Gourley C., Lambrechts D., Chenevix-Trench G., Olsson H., Benitez J.J., Greene M.H., Gore M., Nussbaum R., Sadetzki S., Gayther S.A., Kjaer S.K., D'Andrea A.D., Shapiro G.I., Wiest D.L., Connolly D.C., Daly M.B., Swisher E.M., Bouwman P., Jonkers J., Balmana J., Serra V., Johnson N. The BRCA1-delta11q alternative splice isoform bypasses germline mutations and promotes therapeutic resistance to PARP inhibition and cisplatin. Cancer Res. 2016;76(9):2778–2790. - PMC - PubMed
-
- Seo A., Steinberg-Shemer O., Unal S., Casadei S., Walsh T., Gumruk F., Shalev S., Shimamura A., Akarsu N.A., Tamary H., King M.C. vol. 115. 2018. Mechanism for survival of homozygous nonsense mutations in the tumor suppressor gene BRCA1; pp. 5241–5246. (Proceedings of the National Academy of Science of the United States of America). 20. - PMC - PubMed
-
- Gowen L., Johnson B., Latour A., Sulik K., Koller B. Brca1 deficiency results in early embryonic lethality characterized by neuroepithelial abnormalities. Nat. Genet. 1996;12:191–194. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
