

A neuron-glia lipid metabolic cycle couples daily sleep to mitochondrial homeostasis
- PMID: 38360946
- PMCID: PMC11001586
- DOI: 10.1038/s41593-023-01568-1
A neuron-glia lipid metabolic cycle couples daily sleep to mitochondrial homeostasis
Abstract
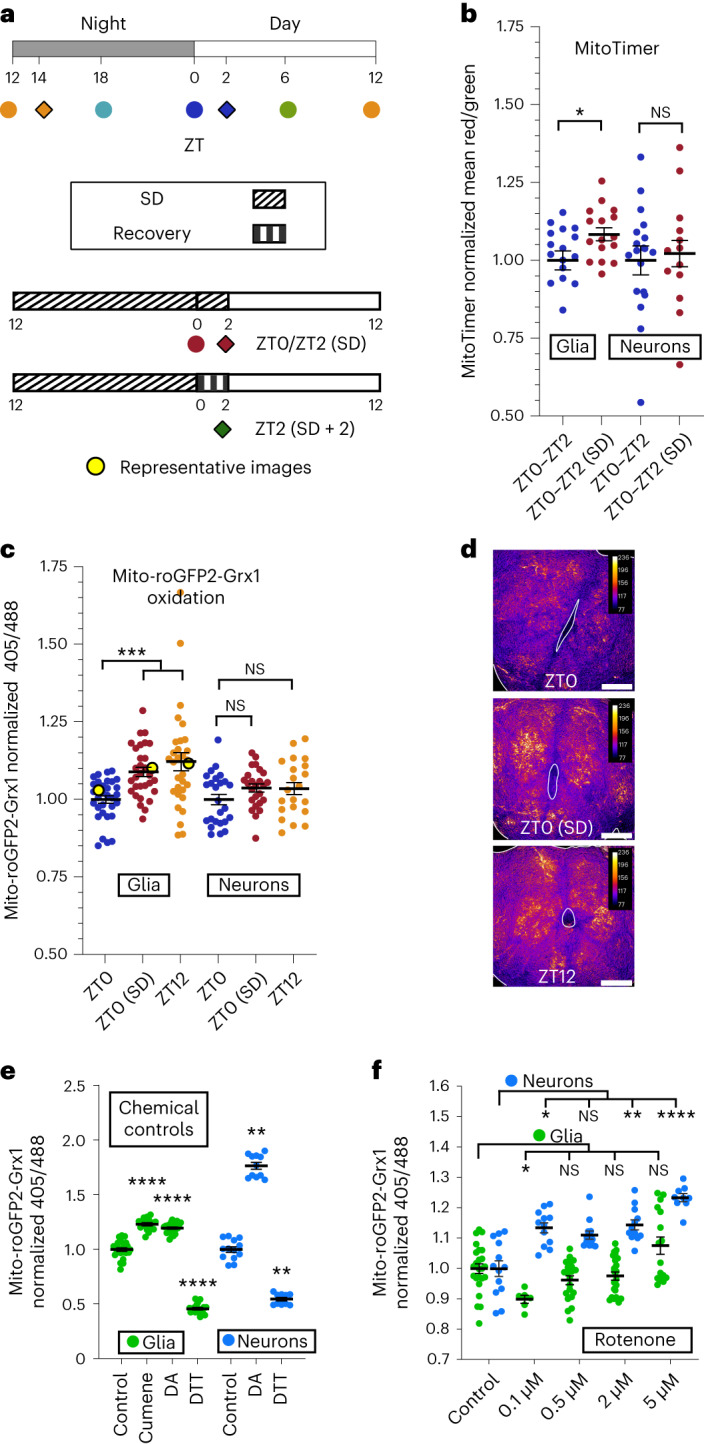
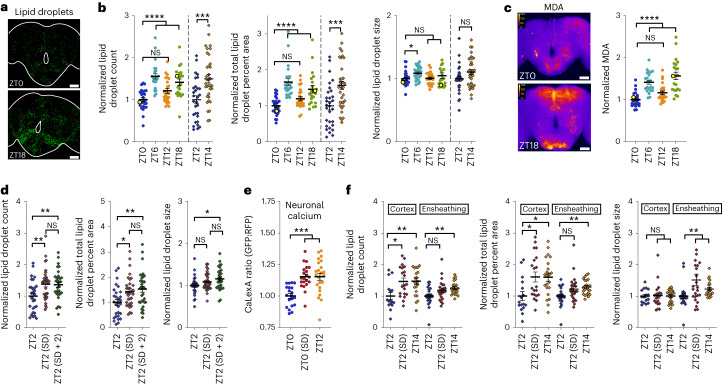
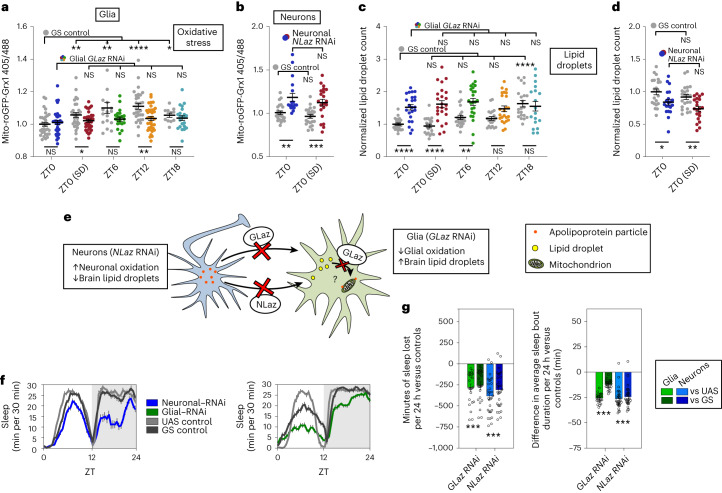
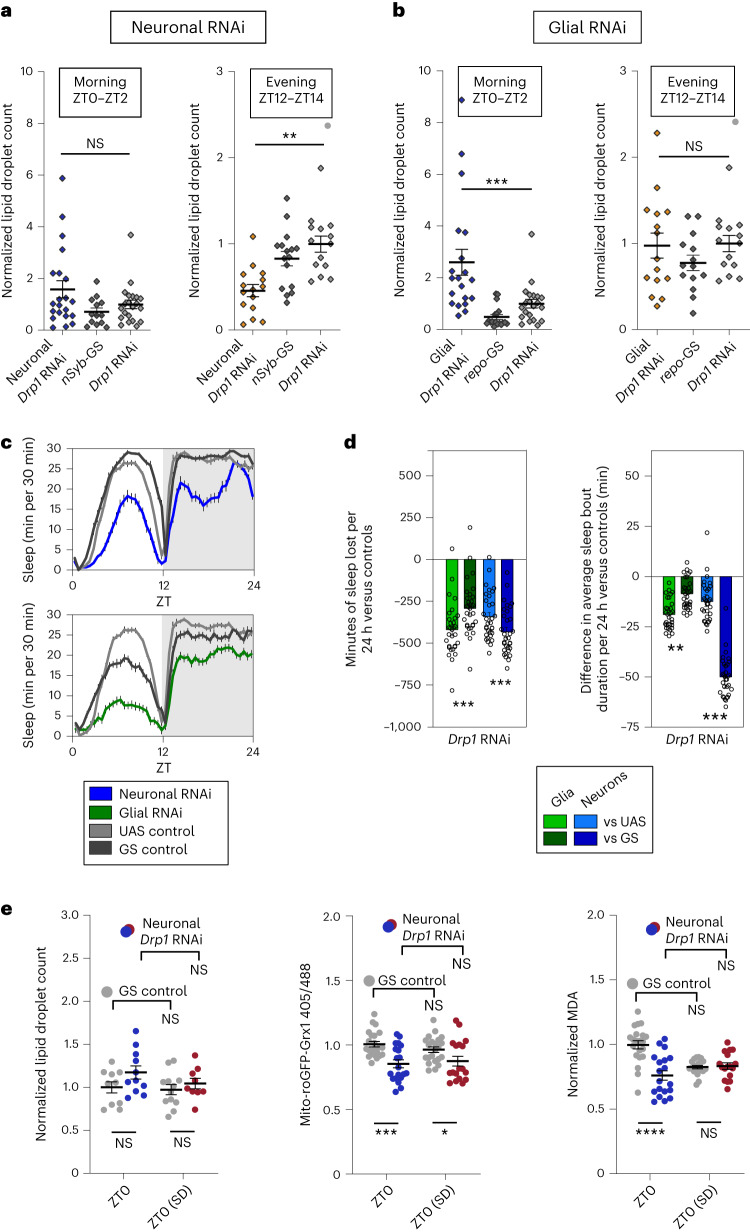
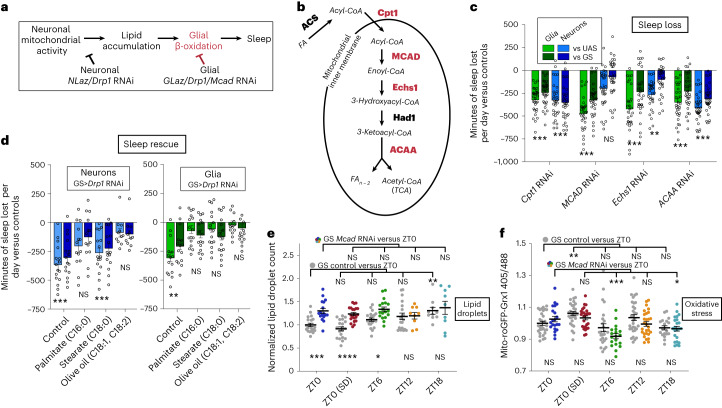
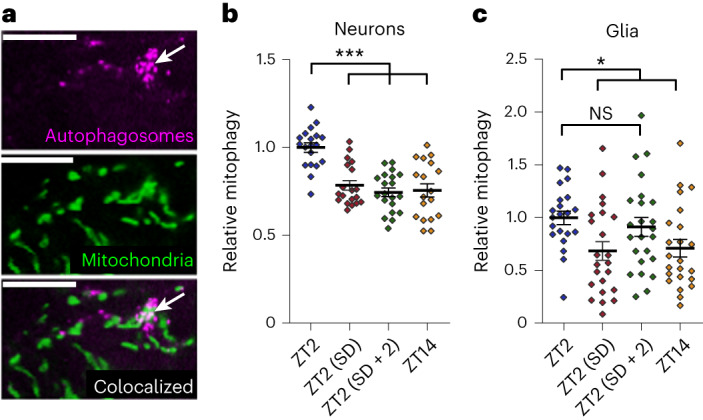
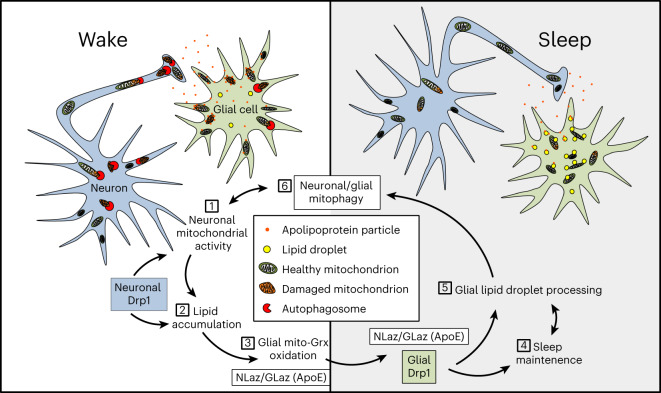
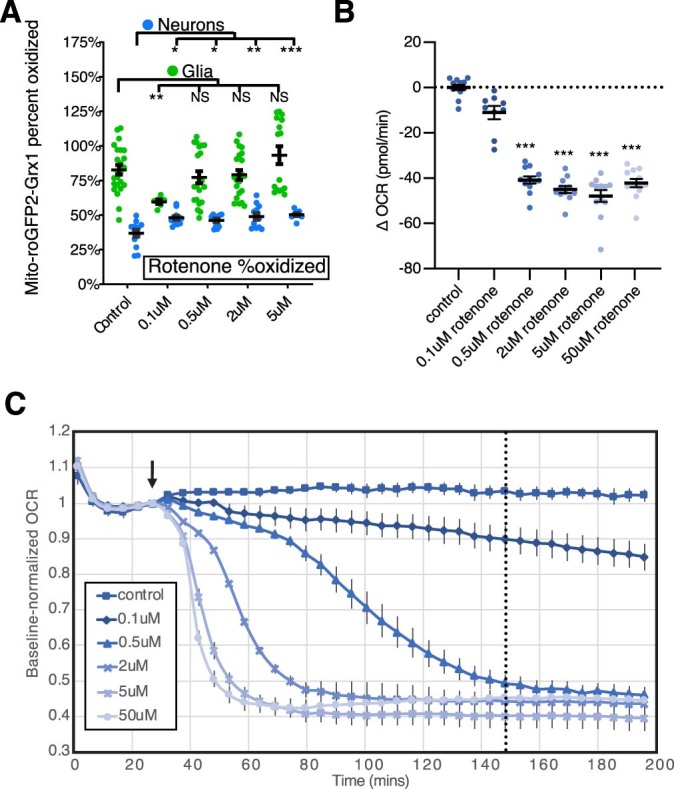
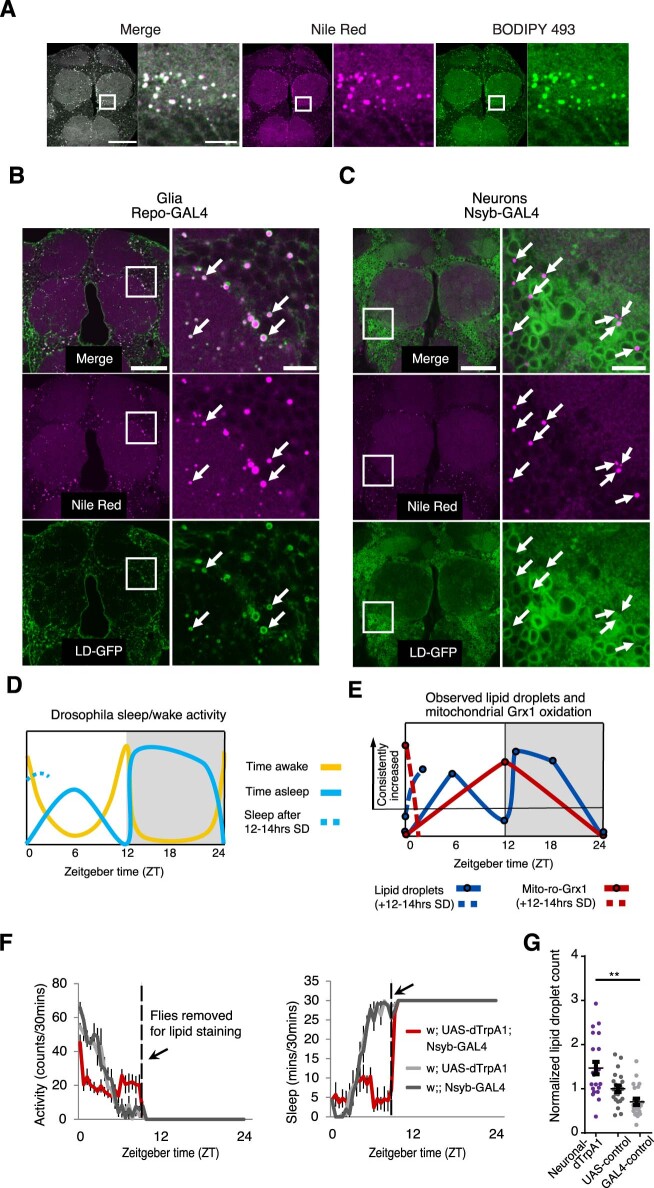
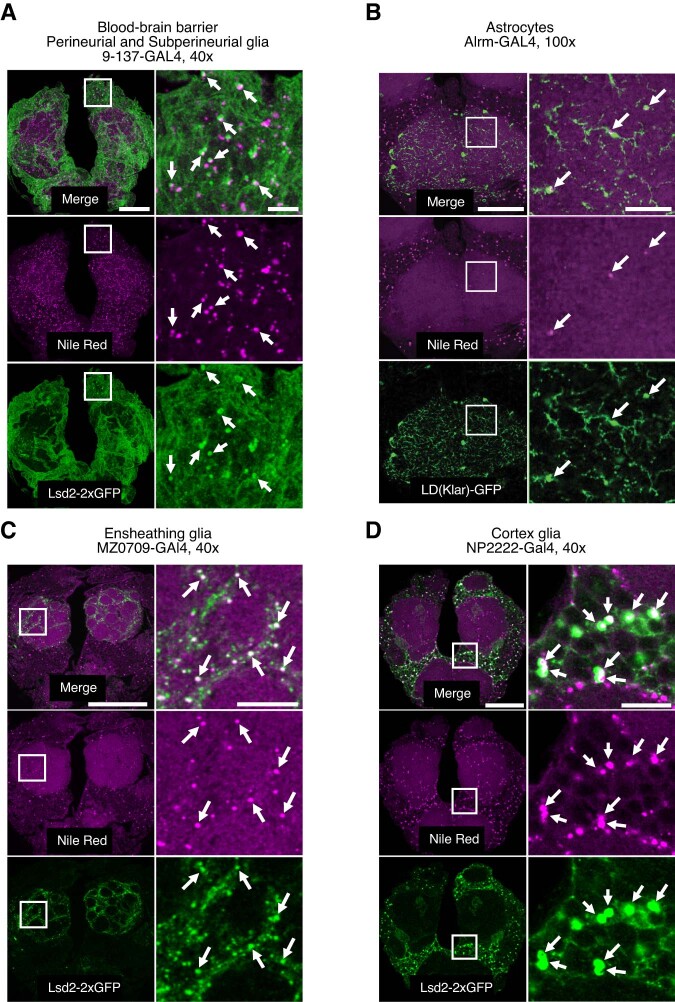
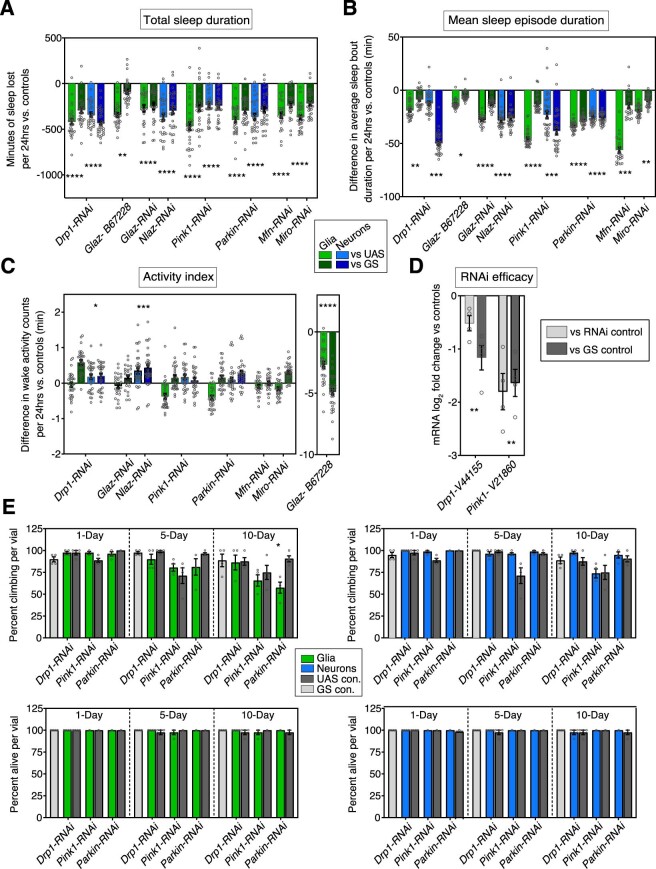
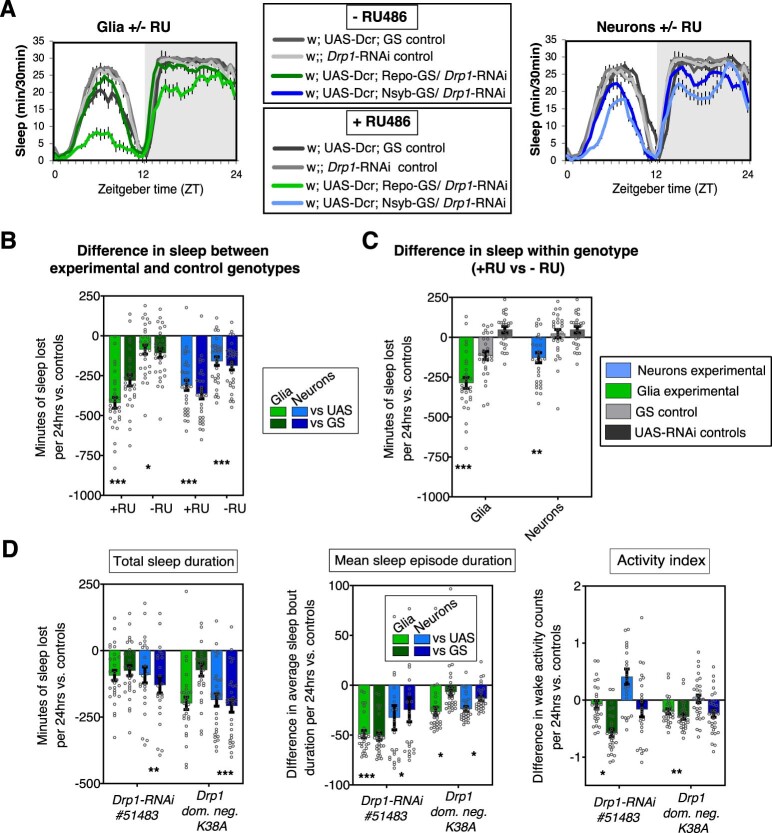
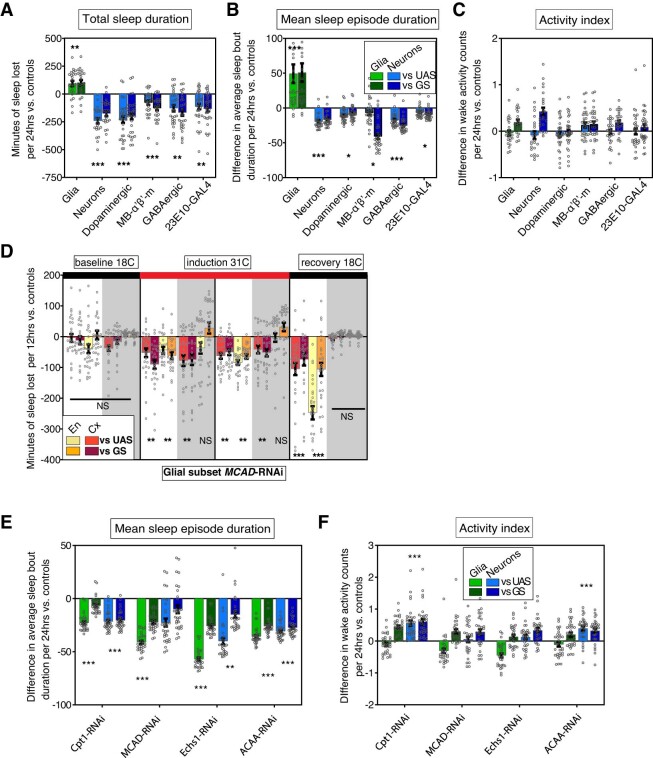
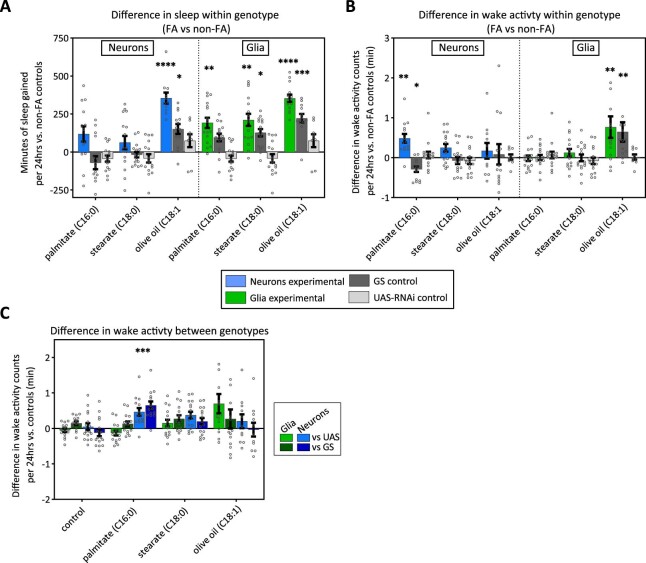
Sleep is thought to be restorative to brain energy homeostasis, but it is not clear how this is achieved. We show here that Drosophila glia exhibit a daily cycle of glial mitochondrial oxidation and lipid accumulation that is dependent on prior wake and requires the Drosophila APOE orthologs NLaz and GLaz, which mediate neuron-glia lipid transfer. In turn, a full night of sleep is required for glial lipid clearance, mitochondrial oxidative recovery and maximal neuronal mitophagy. Knockdown of neuronal NLaz causes oxidative stress to accumulate in neurons, and the neuronal mitochondrial integrity protein, Drp1, is required for daily glial lipid accumulation. These data suggest that neurons avoid accumulation of oxidative mitochondrial damage during wake by using mitophagy and passing damage to glia in the form of lipids. We propose that a mitochondrial lipid metabolic cycle between neurons and glia reflects a fundamental function of sleep relevant for brain energy homeostasis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
