

Aldehyde-mediated inhibition of asparagine biosynthesis has implications for diabetes and alcoholism
- PMID: 38362406
- PMCID: PMC10866355
- DOI: 10.1039/d3sc06551k
Aldehyde-mediated inhibition of asparagine biosynthesis has implications for diabetes and alcoholism
Abstract
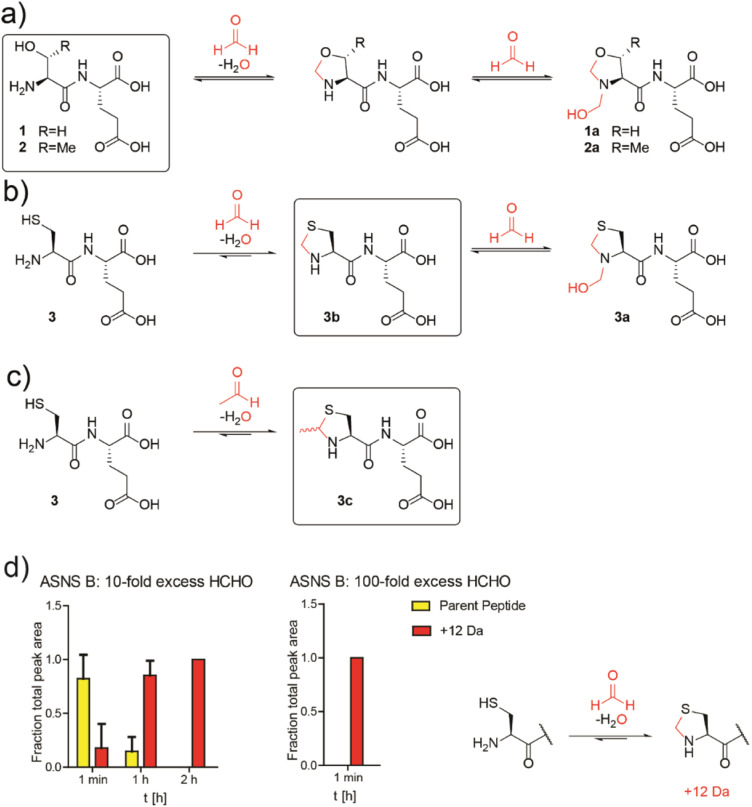
Patients with alcoholism and type 2 diabetes manifest altered metabolism, including elevated aldehyde levels and unusually low asparagine levels. We show that asparagine synthetase B (ASNS), the only human asparagine-forming enzyme, is inhibited by disease-relevant reactive aldehydes, including formaldehyde and acetaldehyde. Cellular studies show non-cytotoxic amounts of reactive aldehydes induce a decrease in asparagine levels. Biochemical analyses reveal inhibition results from reaction of the aldehydes with the catalytically important N-terminal cysteine of ASNS. The combined cellular and biochemical results suggest a possible mechanism underlying the low asparagine levels in alcoholism and diabetes. The results will stimulate research on the biological consequences of the reactions of aldehydes with nucleophilic residues.
This journal is © The Royal Society of Chemistry.
Conflict of interest statement
There are no conflicts to declare.
Figures


References
Grants and funding
LinkOut - more resources
Full Text Sources