

Drosophila melanogaster Sting mediates Coxiella burnetii infection by reducing accumulation of reactive oxygen species
- PMID: 38363133
- PMCID: PMC10929449
- DOI: 10.1128/iai.00560-22
Drosophila melanogaster Sting mediates Coxiella burnetii infection by reducing accumulation of reactive oxygen species
Abstract
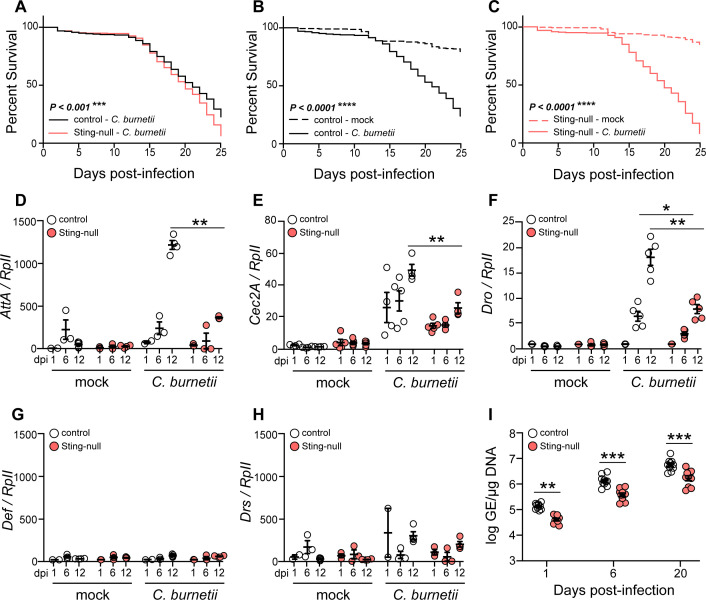
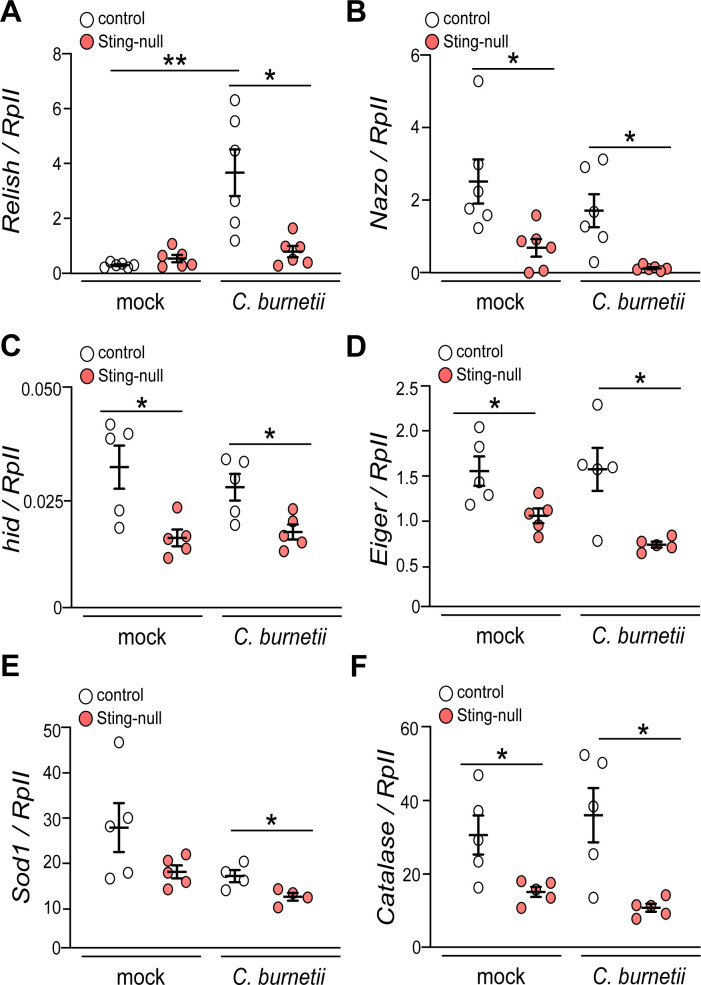
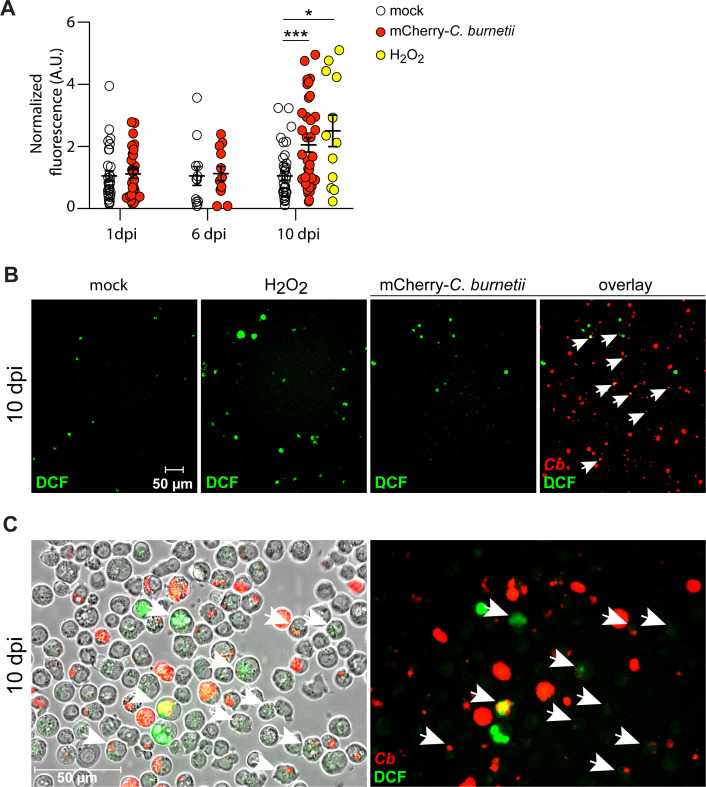
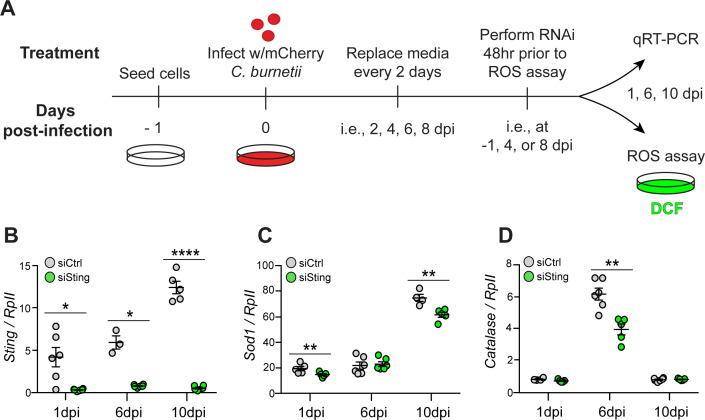
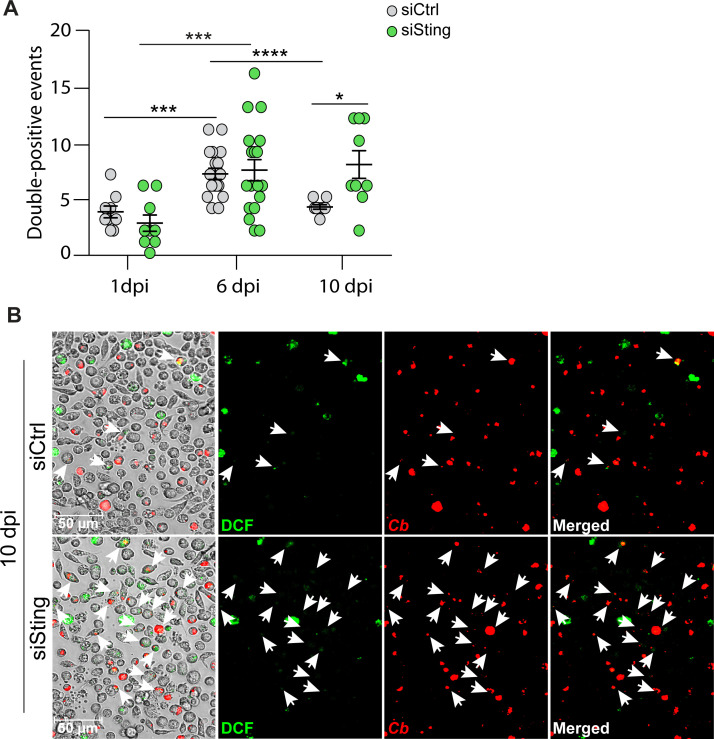
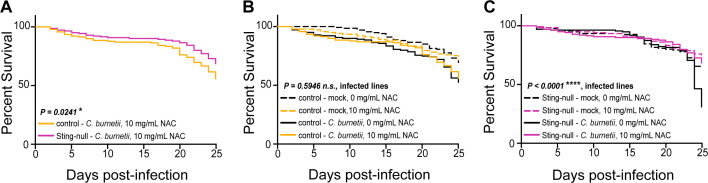
The Gram-negative bacterium Coxiella burnetii is the causative agent of query fever in humans and coxiellosis in livestock. C. burnetii infects a variety of cell types, tissues, and animal species including mammals and arthropods, but there is much left to be understood about the molecular mechanisms at play during infection in distinct species. Human stimulator of interferon genes (STING) induces an innate immune response through the induction of type I interferons (IFNs), and IFN promotes or suppresses C. burnetii replication, depending on tissue type. Drosophila melanogaster contains a functional STING ortholog (Sting) which activates NF-κB signaling and autophagy. Here, we sought to address the role of D. melanogaster Sting during C. burnetii infection to uncover how Sting regulates C. burnetii infection in flies. We show that Sting-null flies exhibit higher mortality and reduced induction of antimicrobial peptides following C. burnetii infection compared to control flies. Additionally, Sting-null flies induce lower levels of oxidative stress genes during infection, but the provision of N-acetyl-cysteine (NAC) in food rescues Sting-null host survival. Lastly, we find that reactive oxygen species levels during C. burnetii infection are higher in Drosophila S2 cells knocked down for Sting compared to control cells. Our results show that at the host level, NAC provides protection against C. burnetii infection in the absence of Sting, thus establishing a role for Sting in protection against oxidative stress during C. burnetii infection.
Keywords: ROS; STING; bacteria; fly; pathogenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
