

Sodium oligomannate alters gut microbiota, reduces cerebral amyloidosis and reactive microglia in a sex-specific manner
- PMID: 38365827
- PMCID: PMC10874048
- DOI: 10.1186/s13024-023-00700-w
Sodium oligomannate alters gut microbiota, reduces cerebral amyloidosis and reactive microglia in a sex-specific manner
Erratum in
-
Correction: Sodium oligomannate alters gut microbiota, reduces cerebral amyloidosis and reactive microglia in a sex-specific manner.Mol Neurodegener. 2024 Oct 10;19(1):70. doi: 10.1186/s13024-024-00764-2. Mol Neurodegener. 2024. PMID: 39390529 Free PMC article. No abstract available.
Abstract
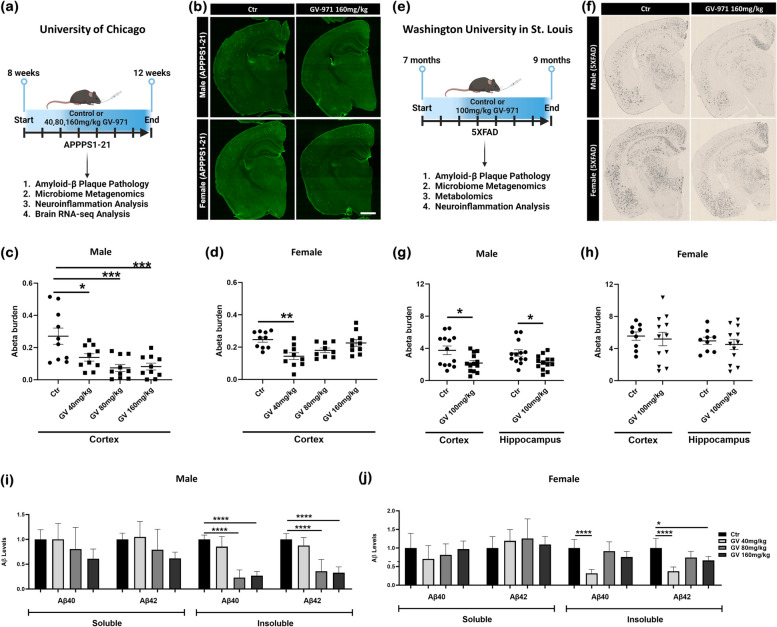
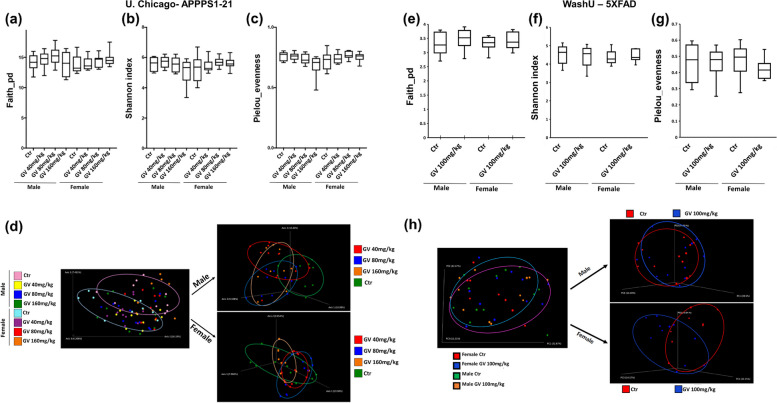
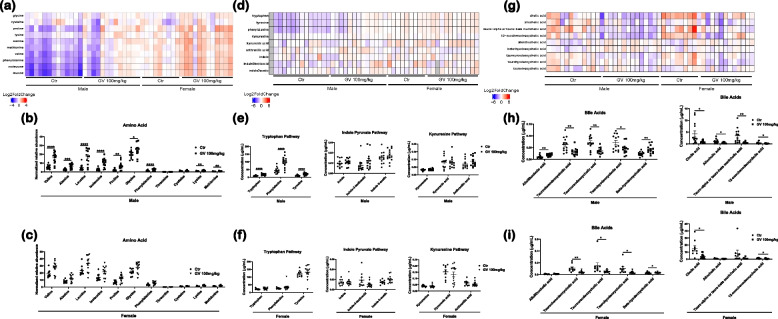
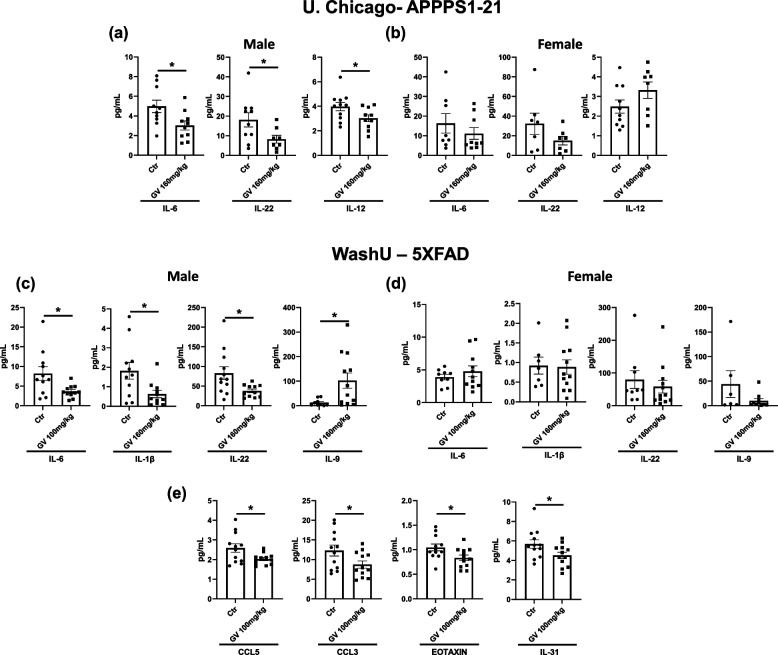
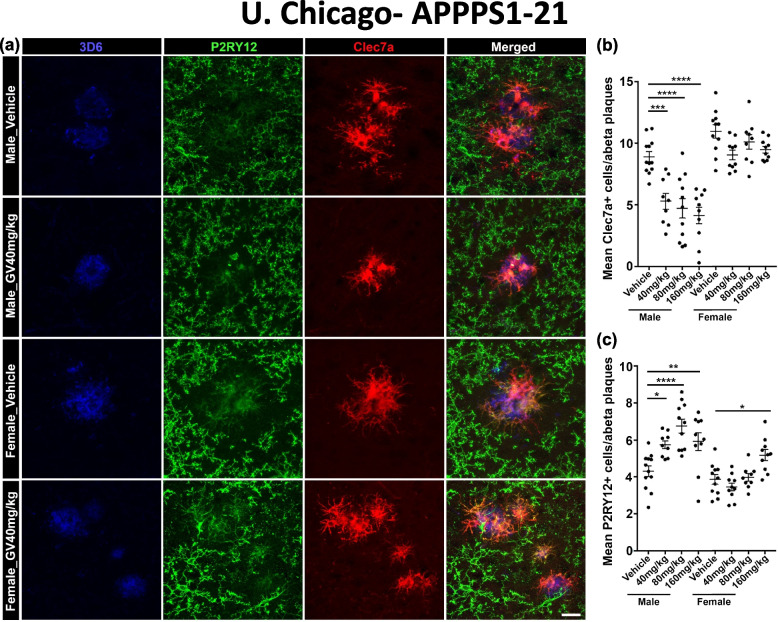
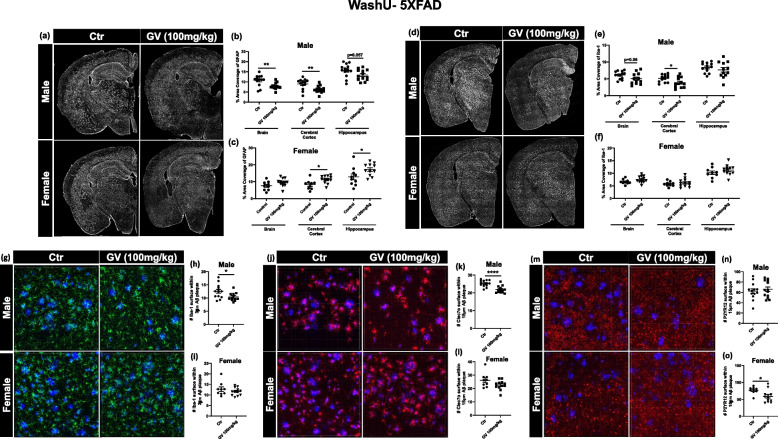
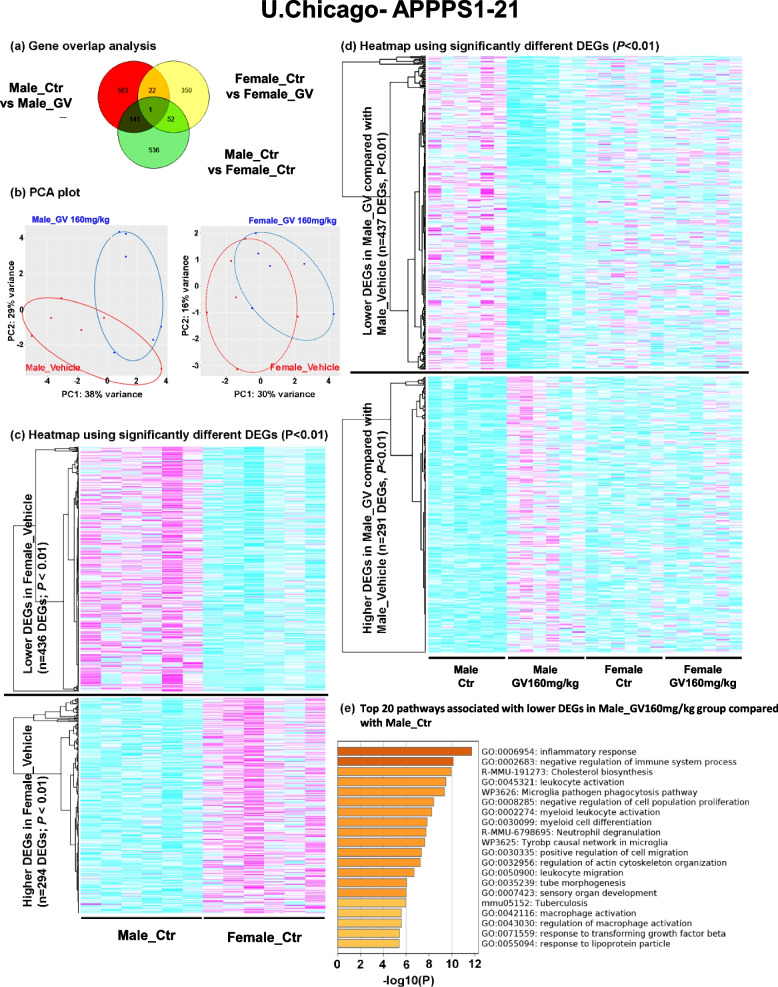
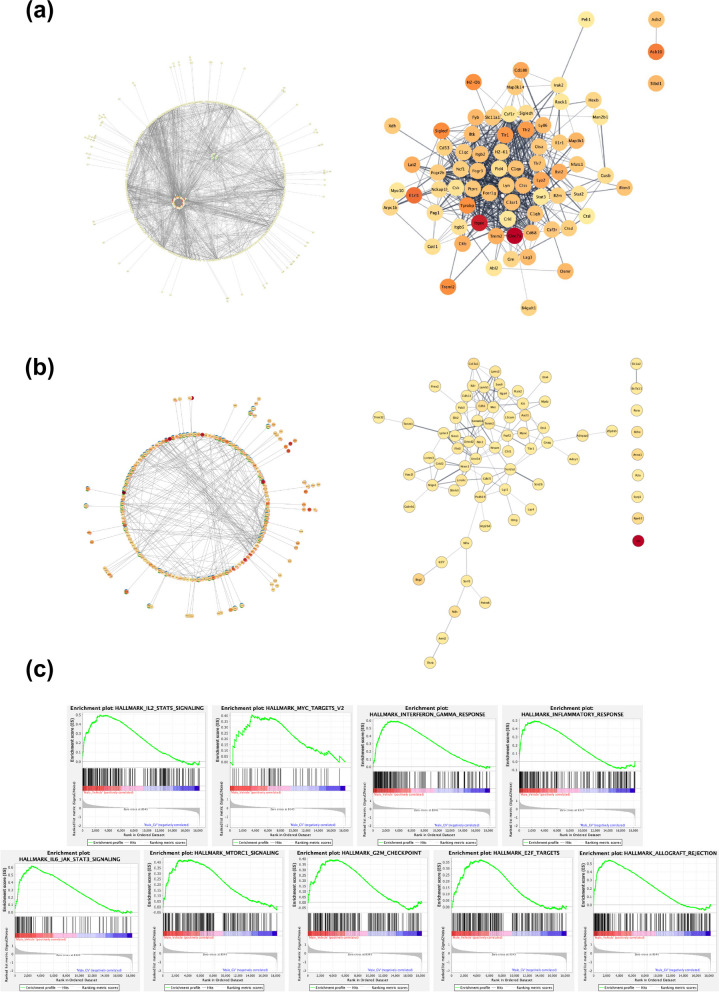
It has recently become well-established that there is a connection between Alzheimer's disease pathology and gut microbiome dysbiosis. We have previously demonstrated that antibiotic-mediated gut microbiota perturbations lead to attenuation of Aβ deposition, phosphorylated tau accumulation, and disease-associated glial cell phenotypes in a sex-dependent manner. In this regard, we were intrigued by the finding that a marine-derived oligosaccharide, GV-971, was reported to alter gut microbiota and reduce Aβ amyloidosis in the 5XFAD mouse model that were treated at a point when Aβ burden was near plateau levels. Utilizing comparable methodologies, but with distinct technical and temporal features, we now report on the impact of GV-971 on gut microbiota, Aβ amyloidosis and microglial phenotypes in the APPPS1-21 model, studies performed at the University of Chicago, and independently in the 5X FAD model, studies performed at Washington University, St. Louis.Methods To comprehensively characterize the effects of GV-971 on the microbiota-microglia-amyloid axis, we conducted two separate investigations at independent institutions. There was no coordination of the experimental design or execution between the two laboratories. Indeed, the two laboratories were not aware of each other's experiments until the studies were completed. Male and female APPPS1-21 mice were treated daily with 40, 80, or 160 mg/kg of GV-971 from 8, when Aβ burden was detectable upto 12 weeks of age when Aβ burden was near maximal levels. In parallel, and to corroborate existing published studies and further investigate sex-related differences, male and female 5XFAD mice were treated daily with 100 mg/kg of GV-971 from 7 to 9 months of age when Aβ burden was near peak levels. Subsequently, the two laboratories independently assessed amyloid-β deposition, metagenomic, and neuroinflammatory profiles. Finally, studies were initiated at the University of Chicago to evaluate the metabolites in cecal tissue from vehicle and GV-971-treated 5XFAD mice.Results These studies showed that independent of the procedural differences (dosage, timing and duration of treatment) between the two laboratories, cerebral amyloidosis was reduced primarily in male mice, independent of strain. We also observed sex-specific microbiota differences following GV-971 treatment. Interestingly, GV-971 significantly altered multiple overlapping bacterial species at both institutions. Moreover, we discovered that GV-971 significantly impacted microbiome metabolism, particularly by elevating amino acid production and influencing the tryptophan pathway. The metagenomics and metabolomics changes correspond with notable reductions in peripheral pro-inflammatory cytokine and chemokine profiles. Furthermore, GV-971 treatment dampened astrocyte and microglia activation, significantly decreasing plaque-associated reactive microglia while concurrently increasing homeostatic microglia only in male mice. Bulk RNAseq analysis unveiled sex-specific changes in cerebral cortex transcriptome profiles, but most importantly, the transcriptome changes in the GV-971-treated male group revealed the involvement of microglia and inflammatory responses.Conclusions In conclusion, these studies demonstrate the connection between the gut microbiome, neuroinflammation, and Alzheimer's disease pathology while highlighting the potential therapeutic effect of GV-971. GV-971 targets the microbiota-microglia-amyloid axis, leading to the lowering of plaque pathology and neuroinflammatory signatures in a sex-dependent manner when given at the onset of Aβ deposition or when given after Aβ deposition is already at higher levels.
Keywords: Alzheimer’s disease; Microbiome; Microglia; Neuroinflammation; Sodium oligomannate.
© 2024. The Author(s).
Conflict of interest statement
D.M.H. co-founded and is on the scientific advisory board of C2N Diagnostics with equity. D.M.H. is on the scientific advisory board of Denali, Genentech, and Cajal Neuroscience and consults for Asteroid. S.S.S. is a paid consultant for Shanghai Green Valley Pharmaceutical Co, Inc.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
