

Modulation of central synapse remodeling after remote peripheral injuries by the CCL2-CCR2 axis and microglia
- PMID: 38367237
- PMCID: PMC10947500
- DOI: 10.1016/j.celrep.2024.113776
Modulation of central synapse remodeling after remote peripheral injuries by the CCL2-CCR2 axis and microglia
Abstract
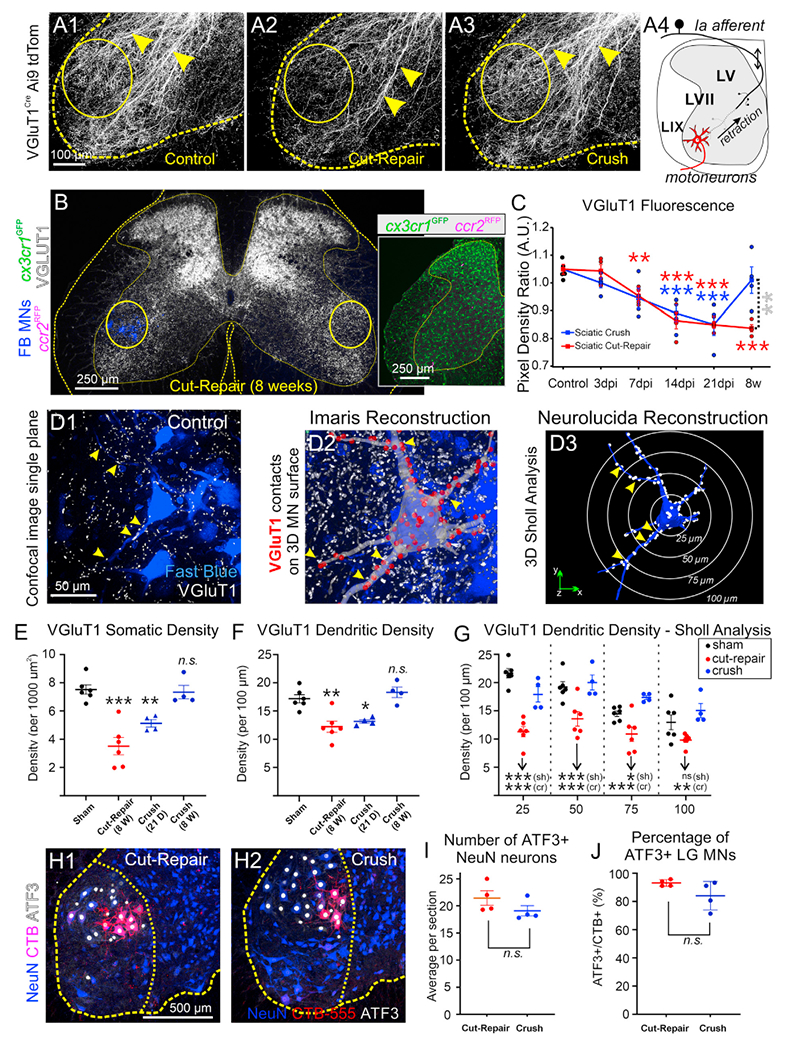
Microglia-mediated synaptic plasticity after CNS injury varies depending on injury severity, but the mechanisms that adjust synaptic plasticity according to injury differences are largely unknown. This study investigates differential actions of microglia on essential spinal motor synaptic circuits following different kinds of nerve injuries. Following nerve transection, microglia and C-C chemokine receptor type 2 signaling permanently remove Ia axons and synapses from the ventral horn, degrading proprioceptive feedback during motor actions and abolishing stretch reflexes. However, Ia synapses and reflexes recover after milder injuries (nerve crush). These different outcomes are related to the length of microglia activation, being longer after nerve cuts, with slower motor-axon regeneration and extended expression of colony-stimulating factor type 1 in injured motoneurons. Prolonged microglia activation induces CCL2 expression, and Ia synapses recover after ccl2 is deleted from microglia. Thus, microglia Ia synapse removal requires the induction of specific microglia phenotypes modulated by nerve regeneration efficiencies. However, synapse preservation was not sufficient to restore the stretch-reflex function.
Keywords: C-C chemokine; CP: Immunology; CP: Neuroscience; Ia afferent; colony stimulating factor 1; motoneuron; nerve injury; neuroinflammation; regeneration; stretch reflex; synapse.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors have no competing interests.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
