

Role of PATJ in stroke prognosis by modulating endothelial to mesenchymal transition through the Hippo/Notch/PI3K axis
- PMID: 38368420
- PMCID: PMC10874379
- DOI: 10.1038/s41420-024-01857-z
Role of PATJ in stroke prognosis by modulating endothelial to mesenchymal transition through the Hippo/Notch/PI3K axis
Abstract
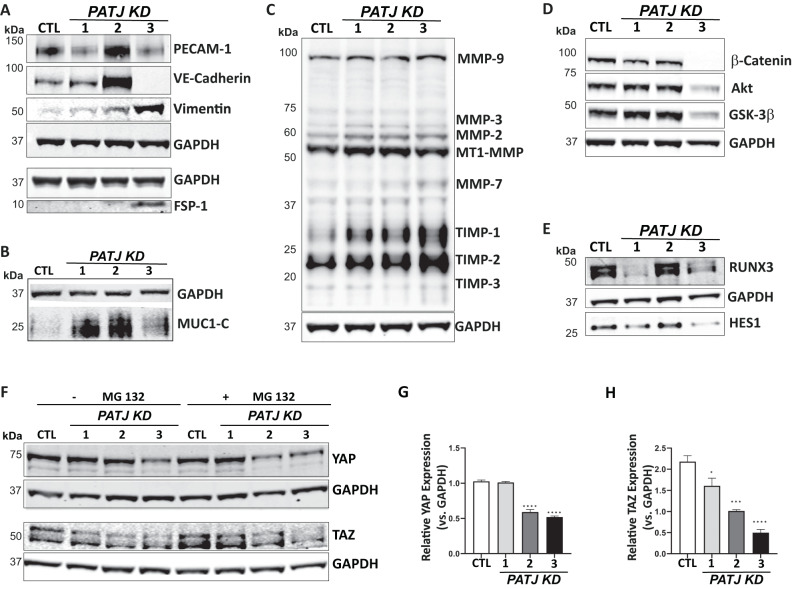
Through GWAS studies we identified PATJ associated with functional outcome after ischemic stroke (IS). The aim of this study was to determine PATJ role in brain endothelial cells (ECs) in the context of stroke outcome. PATJ expression analyses in patient's blood revealed that: (i) the risk allele of rs76221407 induces higher expression of PATJ, (ii) PATJ is downregulated 24 h after IS, and (iii) its expression is significantly lower in those patients with functional independence, measured at 3 months with the modified Rankin scale ((mRS) ≤2), compared to those patients with marked disability (mRS = 4-5). In mice brains, PATJ was also downregulated in the injured hemisphere at 48 h after ischemia. Oxygen-glucose deprivation and hypoxia-dependent of Hypoxia Inducible Factor-1α also caused PATJ depletion in ECs. To study the effects of PATJ downregulation, we generated PATJ-knockdown human microvascular ECs. Their transcriptomic profile evidenced a complex cell reprogramming involving Notch, TGF-ß, PI3K/Akt, and Hippo signaling that translates in morphological and functional changes compatible with endothelial to mesenchymal transition (EndMT). PATJ depletion caused loss of cell-cell adhesion, upregulation of metalloproteases, actin cytoskeleton remodeling, cytoplasmic accumulation of the signal transducer C-terminal transmembrane Mucin 1 (MUC1-C) and downregulation of Notch and Hippo signaling. The EndMT phenotype of PATJ-depleted cells was associated with the nuclear recruitment of MUC1-C, YAP/TAZ, β-catenin, and ZEB1. Our results suggest that PATJ downregulation 24 h after IS promotes EndMT, an initial step prior to secondary activation of a pro-angiogenic program. This effect is associated with functional independence suggesting that activation of EndMT shortly after stroke onset is beneficial for stroke recovery.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






References
Grants and funding
- PI21/00890/Ministry of Economy and Competitiveness | Instituto de Salud Carlos III (Institute of Health Carlos III)
- PI21/00890/Ministry of Economy and Competitiveness | Instituto de Salud Carlos III (Institute of Health Carlos III)
- PI21/00890/Ministry of Economy and Competitiveness | Instituto de Salud Carlos III (Institute of Health Carlos III)
- PI21/00890/Ministry of Economy and Competitiveness | Instituto de Salud Carlos III (Institute of Health Carlos III)
- PI21/00890/Ministry of Economy and Competitiveness | Instituto de Salud Carlos III (Institute of Health Carlos III)
- PI21/00890/Ministry of Economy and Competitiveness | Instituto de Salud Carlos III (Institute of Health Carlos III)
- PI21/00890/Ministry of Economy and Competitiveness | Instituto de Salud Carlos III (Institute of Health Carlos III)
- PI21/00890/Ministry of Economy and Competitiveness | Instituto de Salud Carlos III (Institute of Health Carlos III)
- 776/C/2011/Fundació la Marató de TV3 (TV3 Marathon Foundation)
- 776/C/2011/Fundació la Marató de TV3 (TV3 Marathon Foundation)
- 776/C/2011/Fundació la Marató de TV3 (TV3 Marathon Foundation)
- 776/C/2011/Fundació la Marató de TV3 (TV3 Marathon Foundation)
- 776/C/2011/Fundació la Marató de TV3 (TV3 Marathon Foundation)
- 776/C/2011/Fundació la Marató de TV3 (TV3 Marathon Foundation)
- 188/C/2017/Fundació la Marató de TV3 (TV3 Marathon Foundation)
- PERIS-SLT017/20/000197/Generalitat de Catalunya (Government of Catalonia)
- CSC-201706170048/China Scholarship Council (CSC)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
