

ALLO-1- and IKKE-1-dependent positive feedback mechanism promotes the initiation of paternal mitochondrial autophagy
- PMID: 38368448
- PMCID: PMC10874384
- DOI: 10.1038/s41467-024-45863-2
ALLO-1- and IKKE-1-dependent positive feedback mechanism promotes the initiation of paternal mitochondrial autophagy
Abstract
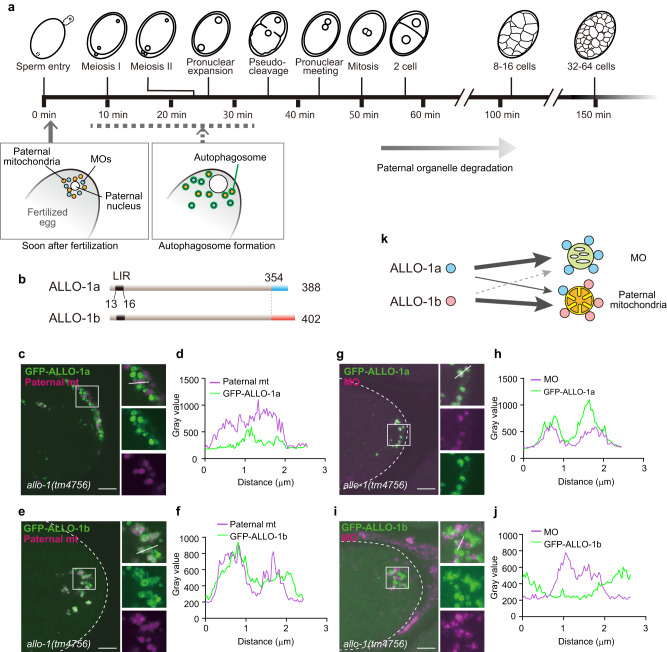
Allophagy is responsible for the selective removal of paternally inherited organelles, including mitochondria, in Caenorhabditis elegans embryos, thereby facilitating the maternal inheritance of mitochondrial DNA. We previously identified two key factors in allophagy: an autophagy adaptor allophagy-1 (ALLO-1) and TBK1/IKKε family kinase IKKE-1. However, the precise mechanisms by which ALLO-1 and IKKE-1 regulate local autophagosome formation remain unclear. In this study, we identify two ALLO-1 isoforms with different substrate preferences during allophagy. Live imaging reveals a stepwise mechanism of ALLO-1 localization with rapid cargo recognition, followed by ALLO-1 accumulation around the cargo. In the ikke-1 mutant, the accumulation of ALLO-1, and not the recognition of cargo, is impaired, resulting in the failure of isolation membrane formation. Our results also suggest a feedback mechanism for ALLO-1 accumulation via EPG-7/ATG-11, a worm homolog of FIP200, which is a candidate for IKKE-1-dependent phosphorylation. This feedback mechanism may underlie the ALLO-1-dependent initiation and progression of autophagosome formation around paternal organelles.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures







References
-
- Sato K, Sato M. Multiple ways to prevent transmission of paternal mitochondrial DNA for maternal inheritance in animals. J. Biochem. 2017;162:247–253. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
