

Polar accumulation of pyoverdin and exit from stationary phase
- PMID: 38370141
- PMCID: PMC10873284
- DOI: 10.1093/femsml/uqae001
Polar accumulation of pyoverdin and exit from stationary phase
Abstract
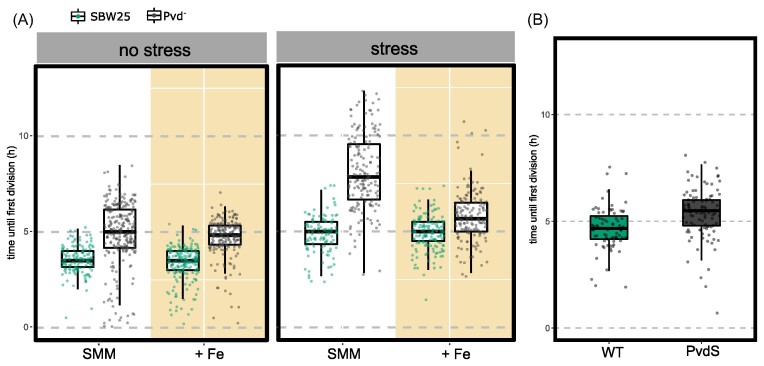
Pyoverdin is a water-soluble metal-chelator synthesized by members of the genus Pseudomonas and used for the acquisition of insoluble ferric iron. Although freely diffusible in aqueous environments, preferential dissemination of pyoverdin among adjacent cells, fine-tuning of intracellular siderophore concentrations, and fitness advantages to pyoverdin-producing versus nonproducing cells, indicate control of location and release. Here, using time-lapse fluorescence microscopy to track single cells in growing microcolonies of Pseudomonas fluorescens SBW25, we show accumulation of pyoverdin at cell poles. Accumulation occurs on cessation of cell growth, is achieved by cross-feeding in pyoverdin-nonproducing mutants and is reversible. Moreover, accumulation coincides with localization of a fluorescent periplasmic reporter, suggesting that pyoverdin accumulation at cell poles is part of the general cellular response to starvation. Compatible with this conclusion is absence of non-accumulating phenotypes in a range of pyoverdin mutants. Analysis of the performance of pyoverdin-producing and nonproducing cells under conditions promoting polar accumulation shows an advantage to accumulation on resumption of growth after stress. Examination of pyoverdin polar accumulation in a multispecies community and in a range of laboratory and natural species of Pseudomonas, including P. aeruginosa PAO1 and P. putida KT2440, confirms that the phenotype is characteristic of Pseudomonas.
Keywords: Pseudomonas; accumulation; imaging; iron; polarization; pyoverdin.
© The Author(s) 2024. Published by Oxford University Press on behalf of FEMS.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Albrecht-Gary AM, Blanc S, Rochel N et al. Bacterial iron transport: coordination properties of pyoverdin PaA, a peptidic siderophore of Pseudomonas aeruginosa. Inorga Chem. 1994;33:6391–402.
LinkOut - more resources
Full Text Sources