

This is a preprint.
T-cells specific for KSHV and HIV migrate to Kaposi sarcoma tumors and persist over time
- PMID: 38370623
- PMCID: PMC10871354
- DOI: 10.1101/2024.02.06.579223
T-cells specific for KSHV and HIV migrate to Kaposi sarcoma tumors and persist over time
Abstract
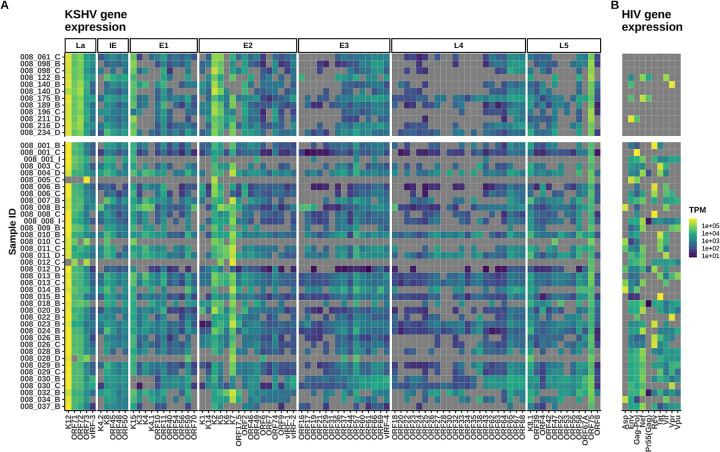
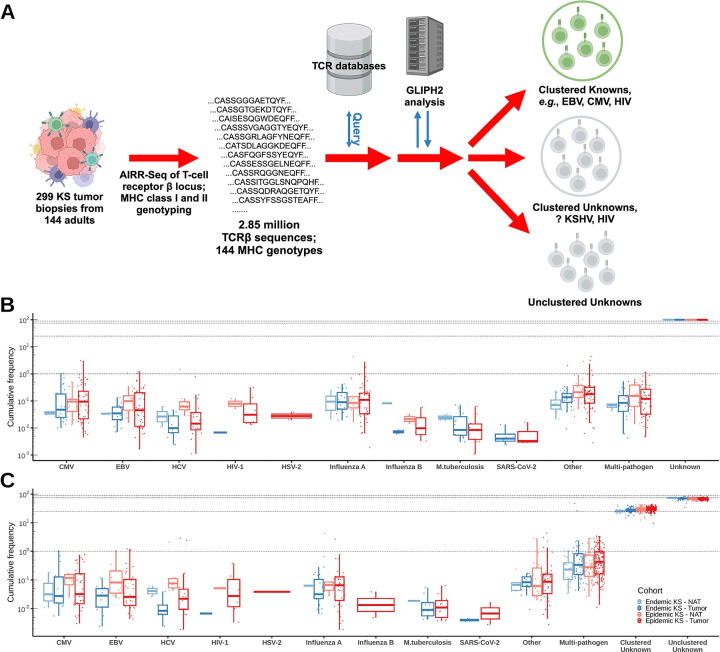
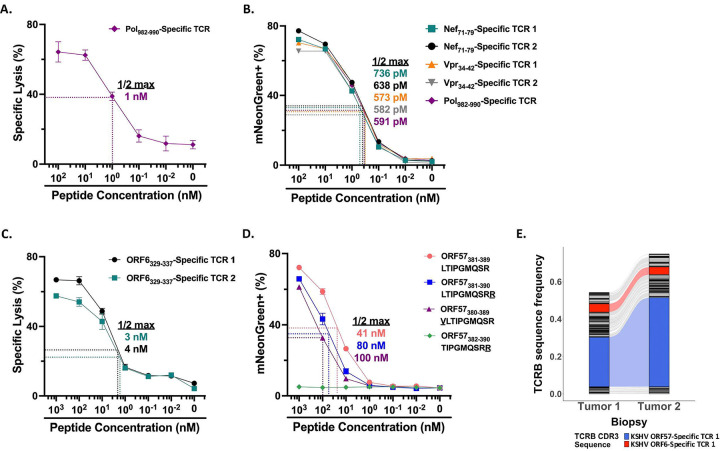
Kaposi sarcoma-associated herpesvirus (KSHV) is the etiologic agent of Kaposi sarcoma (KS), which causes significant morbidity and mortality worldwide, particularly in people living with HIV (PLWH) and in sub-Saharan Africa where KSHV seroprevalence is high. Postulating that T-cells specific for KSHV and HIV would be attracted to KS tumors, we performed transcriptional profiling and T-cell receptor (TCR) repertoire analysis of tumor biopsies from 144 Ugandan adults with KS, 106 of whom were also living with HIV. We show that CD8+ T-cells and M2-polarized macrophages are the most common immune cells in KS tumors. The TCR repertoire of T-cells associated with KS tumors is shared across spatially and temporally distinct tumors from the same individual. Clusters of T-cells with predicted shared specificity for uncharacterized antigens, potentially encoded by KSHV or HIV, comprise ~25% of the T-cells in KS tumors. Single-cell RNA-sequencing of blood from a subset of 9 adults captured 4,283 unique αβ TCRs carried in 14,698 putative KSHV- or HIV-specific T-cells, which carried an antigen-experienced effector phenotype. T-cells engineered to express a representative sample of these TCRs showed high-avidity recognition of KSHV- or HIV-encoded antigens. These results suggest that a poly-specific, high-avidity KSHV- and HIV-specific T-cell response, potentially inhibited by M2 macrophages, migrates to and localizes with KS tumors. Further analysis of KSHV- and HIV-specific T-cells in KS tumors will provide insight into the pathogenesis of KS and could guide the development of specific immune therapy based on adoptive transfer or vaccination.
Conflict of interest statement
Conflict of Interest The authors declare that the research was conducted in the absence of any commercial or financial relationships that could create a conflict of interest.
Figures




Similar articles
-
Indicators for Increased Likelihood of Epidemic Kaposi Sarcoma Progression After Antiretroviral Therapy Initiation.AIDS Res Hum Retroviruses. 2025 Jul;41(7):348-356. doi: 10.1089/aid.2025.0007. Epub 2025 Apr 3. AIDS Res Hum Retroviruses. 2025. PMID: 40178949
-
Comparative polar and lipid plasma metabolomics differentiate KSHV infection and disease states.Cancer Metab. 2023 Aug 31;11(1):13. doi: 10.1186/s40170-023-00316-0. Cancer Metab. 2023. PMID: 37653396 Free PMC article.
-
Single-Cell Transcriptomic Analysis of Kaposi Sarcoma.bioRxiv [Preprint]. 2024 May 3:2024.05.01.592010. doi: 10.1101/2024.05.01.592010. bioRxiv. 2024. Update in: PLoS Pathog. 2025 Apr 1;21(4):e1012233. doi: 10.1371/journal.ppat.1012233. PMID: 38746135 Free PMC article. Updated. Preprint.
-
Treatment of Kaposi sarcoma in children with HIV-1 infection.Cochrane Database Syst Rev. 2014 Jan 27;2014(1):CD009826. doi: 10.1002/14651858.CD009826.pub2. Cochrane Database Syst Rev. 2014. PMID: 24464843 Free PMC article.
-
Antibody tests for identification of current and past infection with SARS-CoV-2.Cochrane Database Syst Rev. 2022 Nov 17;11(11):CD013652. doi: 10.1002/14651858.CD013652.pub2. Cochrane Database Syst Rev. 2022. PMID: 36394900 Free PMC article.
Cited by
-
Single-Cell Transcriptomic Analysis of Kaposi Sarcoma.PLoS Pathog. 2025 Apr 1;21(4):e1012233. doi: 10.1371/journal.ppat.1012233. eCollection 2025 Apr. PLoS Pathog. 2025. PMID: 40168402 Free PMC article.
References
-
- Ibrahim Khalil A, Franceschi S, de Martel C, Bray F, Clifford GM. Burden of Kaposi sarcoma according to HIV status: A systematic review and global analysis. Int J Cancer. 2022;150(12):1948–57. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials