

This is a preprint.
Temporal dynamics of BMP/Nodal ratio drive tissue-specific gastrulation morphogenesis
- PMID: 38370754
- PMCID: PMC10871350
- DOI: 10.1101/2024.02.06.579243
Temporal dynamics of BMP/Nodal ratio drive tissue-specific gastrulation morphogenesis
Update in
-
Temporal dynamics of BMP/Nodal ratio drive tissue-specific gastrulation morphogenesis.Development. 2025 May 1;152(9):dev202931. doi: 10.1242/dev.202931. Epub 2025 May 1. Development. 2025. PMID: 39651654
Abstract
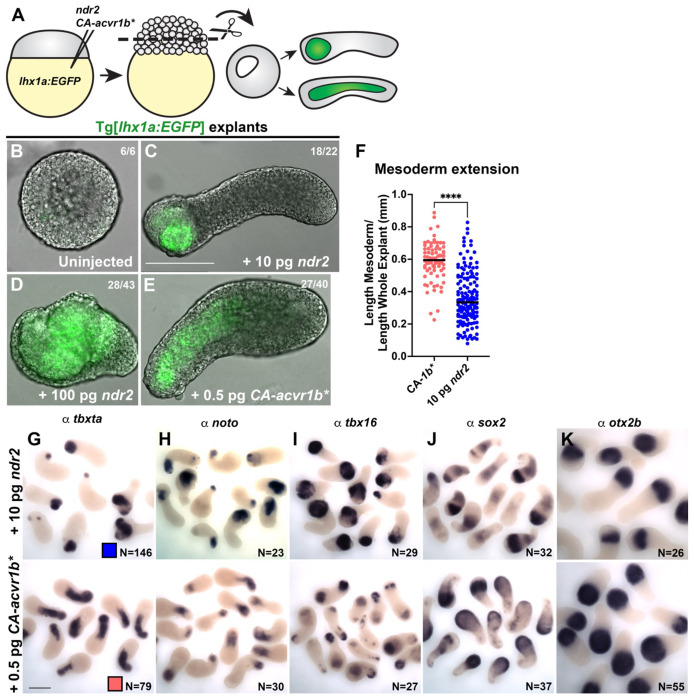
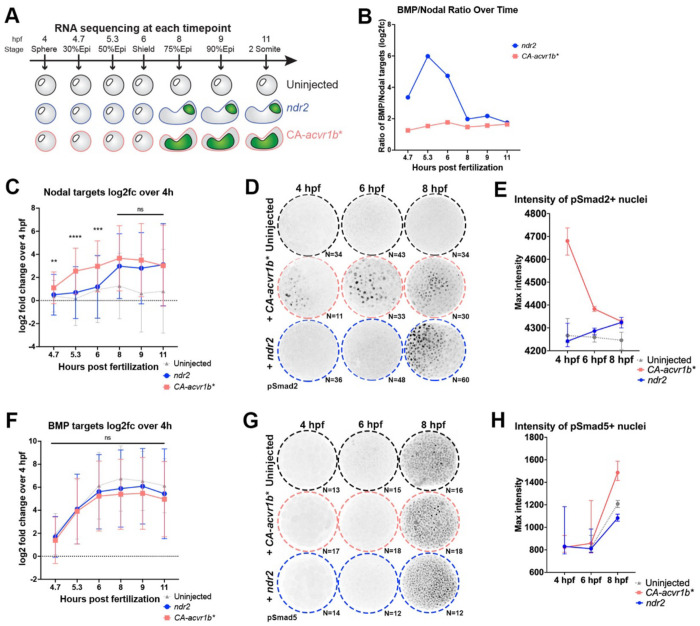
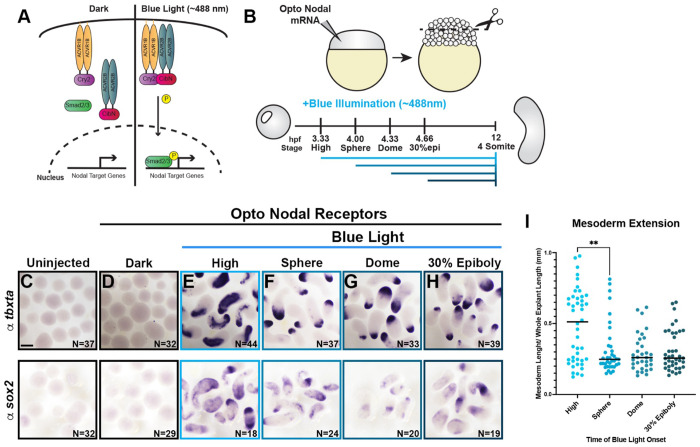
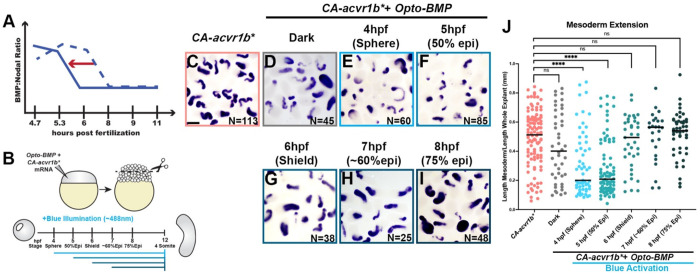
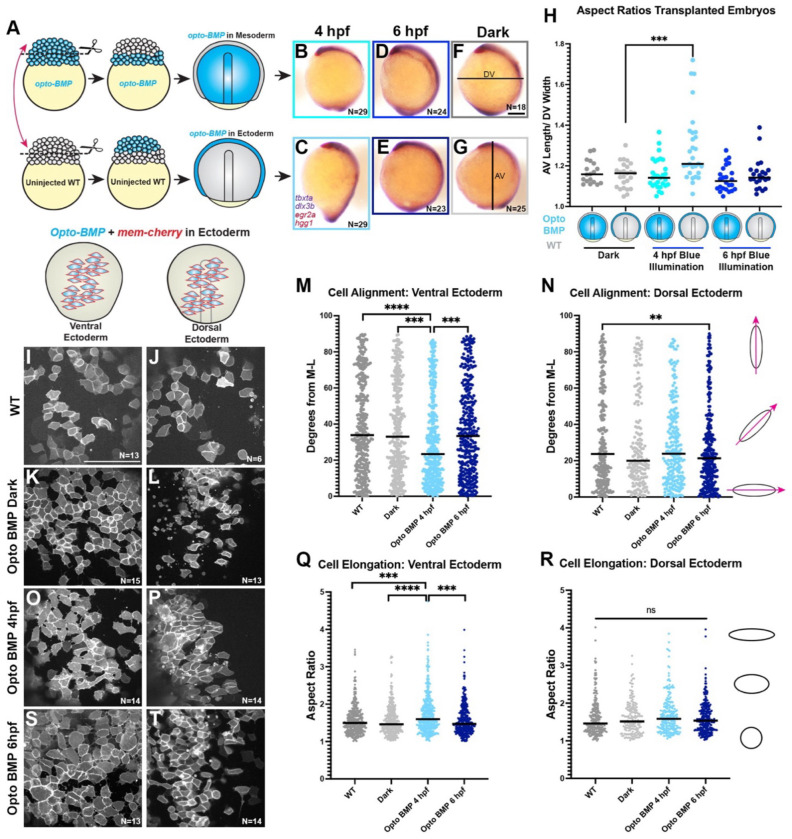
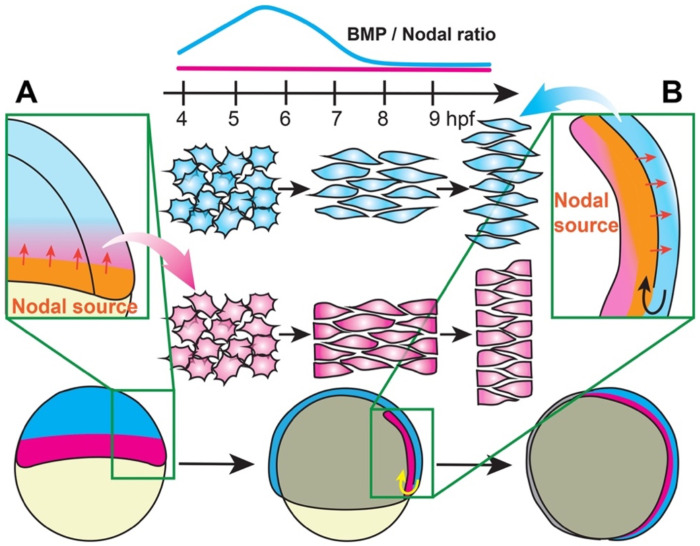
Anteroposterior (AP) elongation of the vertebrate body plan is driven by convergence and extension (C&E) gastrulation movements in both the mesoderm and neuroectoderm, but how or whether molecular regulation of C&E differs between tissues remains an open question. Using a zebrafish explant model of AP axis extension, we show that C&E of the neuroectoderm and mesoderm can be uncoupled ex vivo, and that morphogenesis of individual tissues results from distinct morphogen signaling dynamics. Using precise temporal manipulation of BMP and Nodal signaling, we identify a critical developmental window during which high or low BMP/Nodal ratios induce neuroectoderm- or mesoderm-driven C&E, respectively. Increased BMP activity similarly enhances C&E specifically in the ectoderm of intact zebrafish gastrulae, highlighting the in vivo relevance of our findings. Together, these results demonstrate that temporal dynamics of BMP and Nodal morphogen signaling activate distinct morphogenetic programs governing C&E gastrulation movements within individual tissues.
Keywords: BMP; Convergent extension; Gastrulation; Morphogenesis; Nodal; Zebrafish.
Conflict of interest statement
COMPETING INTERESTS The authors declare no competing interests.
Figures

References
-
- Concha M. L., Adams R. J., Oriented cell divisions and cellular morphogenesis in the zebrafish gastrula and neurula: a time-lapse analysis. Development 125, 983–994 (1998). - PubMed
-
- Warga R. M., Kimmel C. B., Cell movements during epiboly and gastrulation in zebrafish. Development 108, 569–580 (1990). - PubMed
-
- Keller R. E., Danilchik M., Gimlich R., Shih J., The function and mechanism of convergent extension during gastrulation of Xenopus laevis. J Embryol Exp Morphol 89 Suppl, 185–209 (1985). - PubMed
-
- Wallingford J. B., Harland R. M., Xenopus Dishevelled signaling regulates both neural and mesodermal convergent extension: parallel forces elongating the body axis. Development 128, 2581–2592 (2001). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources