

This is a preprint.
Neural signatures of opioid-induced risk-taking behavior in the prelimbic prefrontal cortex
- PMID: 38370807
- PMCID: PMC10871263
- DOI: 10.1101/2024.02.05.578828
Neural signatures of opioid-induced risk-taking behavior in the prelimbic prefrontal cortex
Update in
-
Neural correlates of opioid-induced risk-taking behavior in the prelimbic prefrontal cortex.J Neurosci. 2025 Mar 17;45(19):e2422242025. doi: 10.1523/JNEUROSCI.2422-24.2025. Online ahead of print. J Neurosci. 2025. PMID: 40097184
Abstract
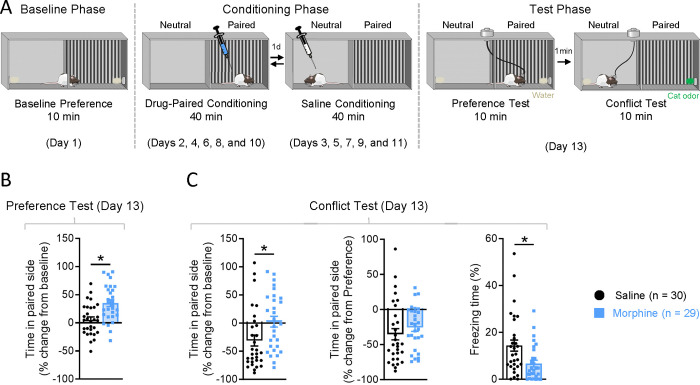
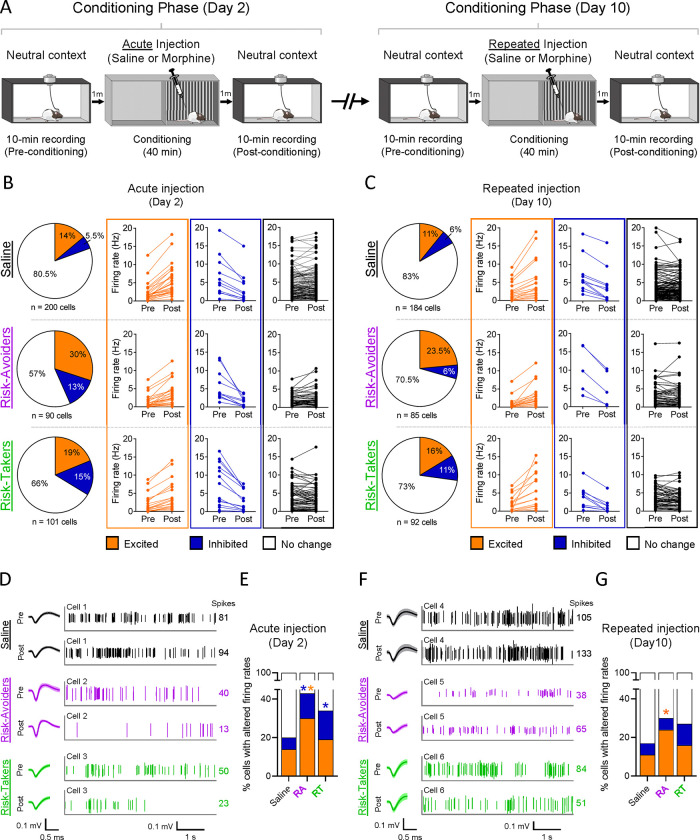
Opioid use disorder occurs alongside impaired risk-related decision-making, but the underlying neural correlates are unclear. We developed an approach-avoidance conflict task using a modified conditioned place preference procedure to study neural signals of risky opioid seeking in the prefrontal cortex, a region implicated in executive decision-making. Following morphine conditioned place preference, rats underwent a conflict test in which fear-inducing cat odor was introduced in the previously drug-paired side of the apparatus. While the saline-exposed control group avoided cat odor, the morphine group included two subsets of rats that either maintained a preference for the paired side despite the presence of cat odor (Risk-Takers) or exhibited increased avoidance (Risk-Avoiders), as revealed by K-means clustering. Single-unit recordings from the prelimbic cortex (PL) demonstrated decreased neuronal activity upon acute morphine exposure in both Risk-Takers and Risk-Avoiders, but this firing rate suppression was absent after repeated morphine administration. Risk-Avoiders also displayed distinct post-morphine excitation in PL which persisted across conditioning. During the preference test, subpopulations of PL neurons in all groups were either excited or inhibited when rats entered the paired side. Interestingly, the inhibition in PL activity was lost during the subsequent conflict test in both saline and Risk-Avoider groups, but persisted in Risk-Takers. Additionally, Risk-Takers showed an increase in the proportion of PL neurons displaying location-specific firing in the drug-paired side from the preference to the conflict test. Together, our results suggest that persistent PL inhibitory signaling in the drug-associated context during motivational conflict may underlie increased risk-taking behavior following opioid exposure.
Conflict of interest statement
COMPETING INTERESTS The authors declare no competing interests.
Figures



Similar articles
-
Neural correlates of opioid-induced risk-taking behavior in the prelimbic prefrontal cortex.J Neurosci. 2025 Mar 17;45(19):e2422242025. doi: 10.1523/JNEUROSCI.2422-24.2025. Online ahead of print. J Neurosci. 2025. PMID: 40097184
-
Neural correlates and determinants of approach-avoidance conflict in the prelimbic prefrontal cortex.Elife. 2021 Dec 16;10:e74950. doi: 10.7554/eLife.74950. Elife. 2021. PMID: 34913438 Free PMC article.
-
Norepinephrine in prelimbic cortex delays extinction of amphetamine-induced conditioned place preference.Psychopharmacology (Berl). 2016 Mar;233(6):973-82. doi: 10.1007/s00213-015-4177-6. Epub 2015 Dec 12. Psychopharmacology (Berl). 2016. PMID: 26660648
-
Temporal Dynamics Underlying Prelimbic Prefrontal Cortical Regulation of Action Selection and Outcome Evaluation during Risk/Reward Decision-Making.J Neurosci. 2023 Feb 15;43(7):1238-1255. doi: 10.1523/JNEUROSCI.0802-22.2022. Epub 2023 Jan 6. J Neurosci. 2023. PMID: 36609453 Free PMC article.
-
Drug-sensitive reward in crayfish: an invertebrate model system for the study of SEEKING, reward, addiction, and withdrawal.Neurosci Biobehav Rev. 2011 Oct;35(9):1847-53. doi: 10.1016/j.neubiorev.2010.12.008. Epub 2010 Dec 21. Neurosci Biobehav Rev. 2011. PMID: 21182861 Free PMC article. Review.
References
-
- Ballesteros-Yáñez I, Ambrosio E, Benavides-Piccione R, Pérez J, Torres I, Miguéns M, García-Lecumberri C, DeFelipe J (2007) The effects of morphine self-administration on cortical pyramidal cell structure in addiction-prone lewis rats. Cereb Cortex N Y N 1991 17:238–249. - PubMed
-
- Bardo MT, Rowlett JK, Harris MJ (1995) Conditioned place preference using opiate and stimulant drugs: A meta-analysis. Neurosci Biobehav Rev 19:39–51. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous