

Metabolic adaptation towards glycolysis supports resistance to neoadjuvant chemotherapy in early triple negative breast cancers
- PMID: 38374113
- PMCID: PMC10875828
- DOI: 10.1186/s13058-024-01788-8
Metabolic adaptation towards glycolysis supports resistance to neoadjuvant chemotherapy in early triple negative breast cancers
Abstract
Background: Neoadjuvant chemotherapy (NAC) is the standard of care for patients with early-stage triple negative breast cancers (TNBC). However, more than half of TNBC patients do not achieve a pathological complete response (pCR) after NAC, and residual cancer burden (RCB) is associated with dismal long-term prognosis. Understanding the mechanisms underlying differential treatment outcomes is therefore critical to limit RCB and improve NAC efficiency.
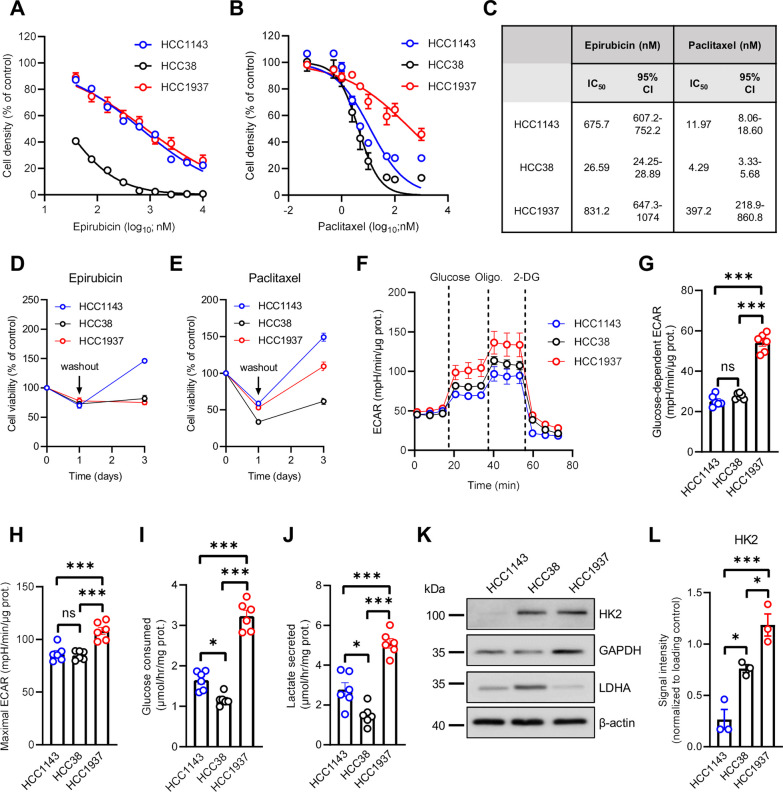
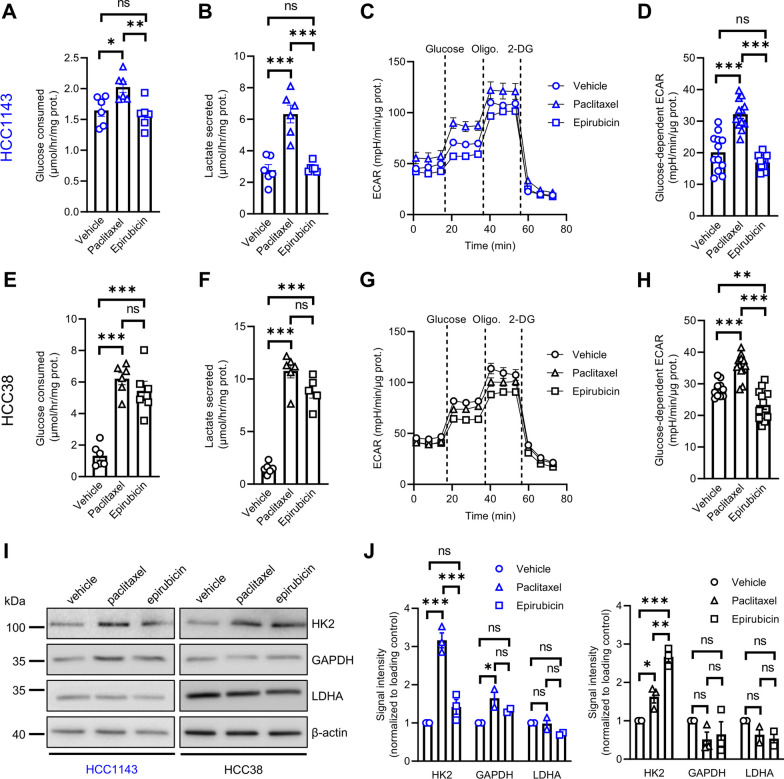
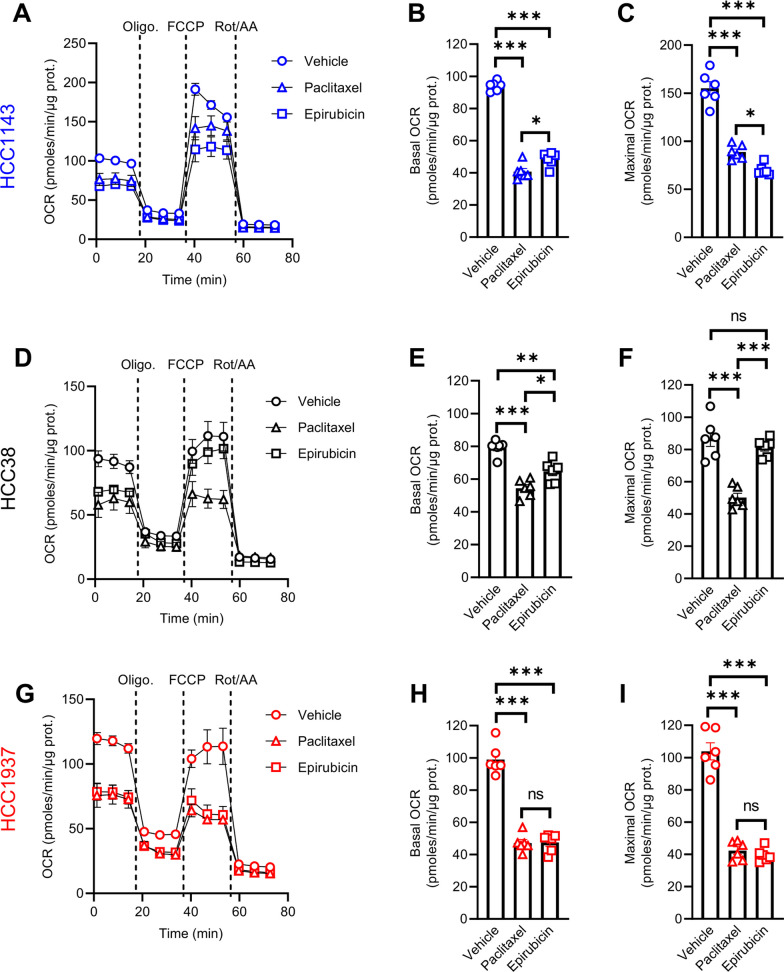
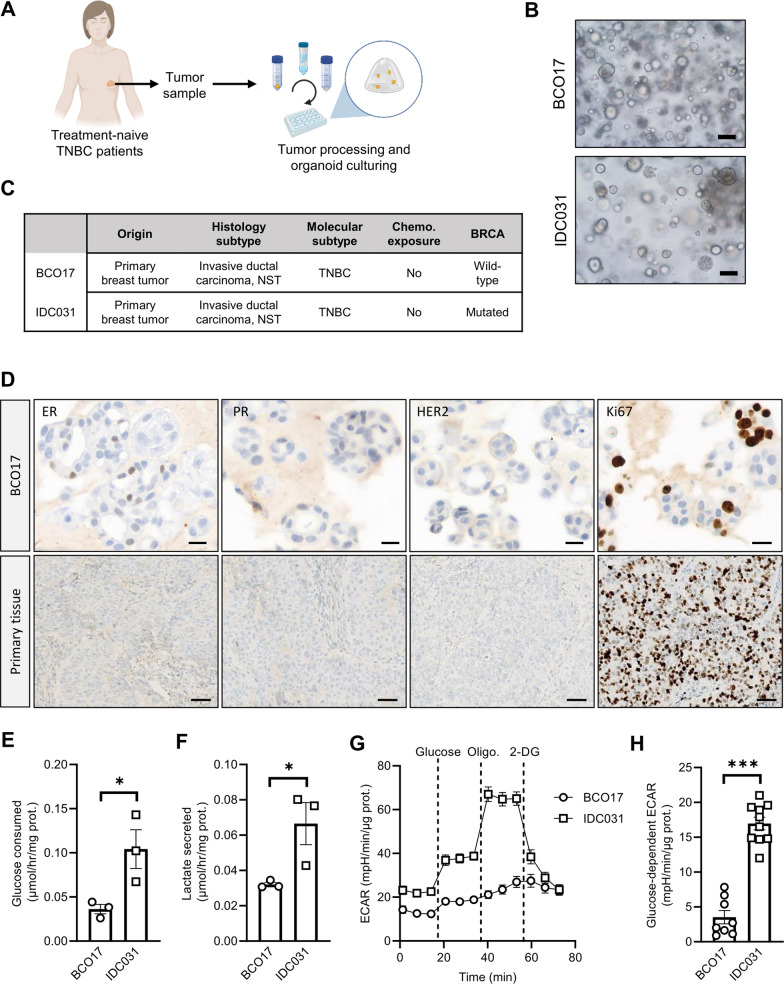
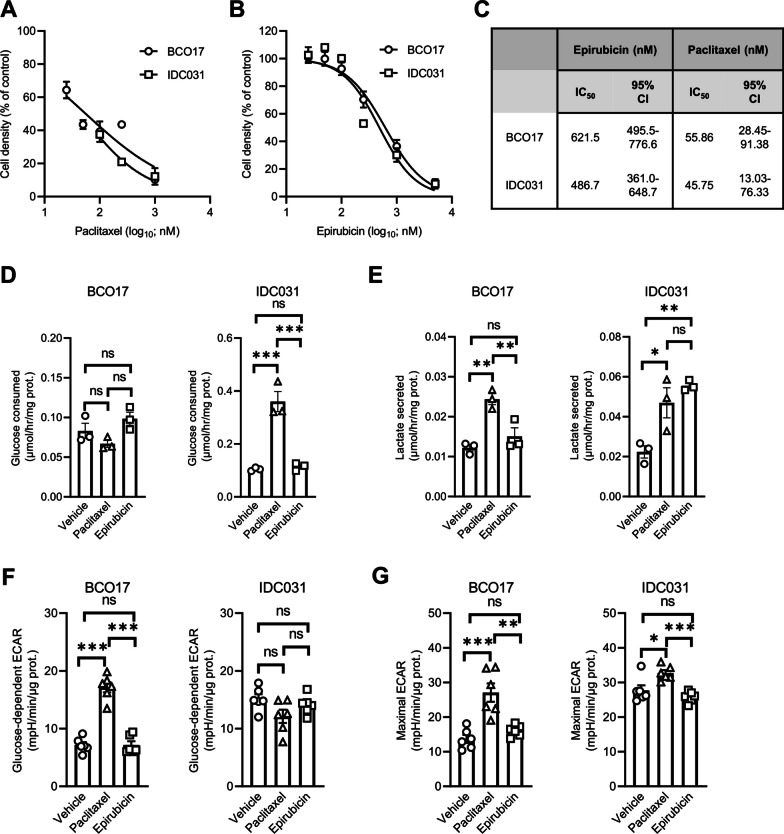
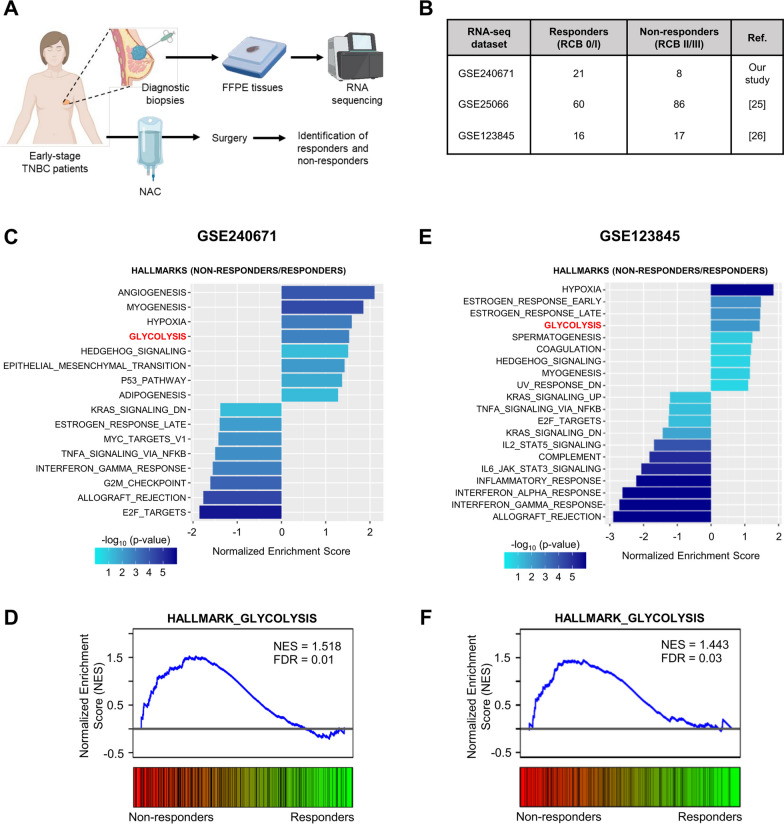
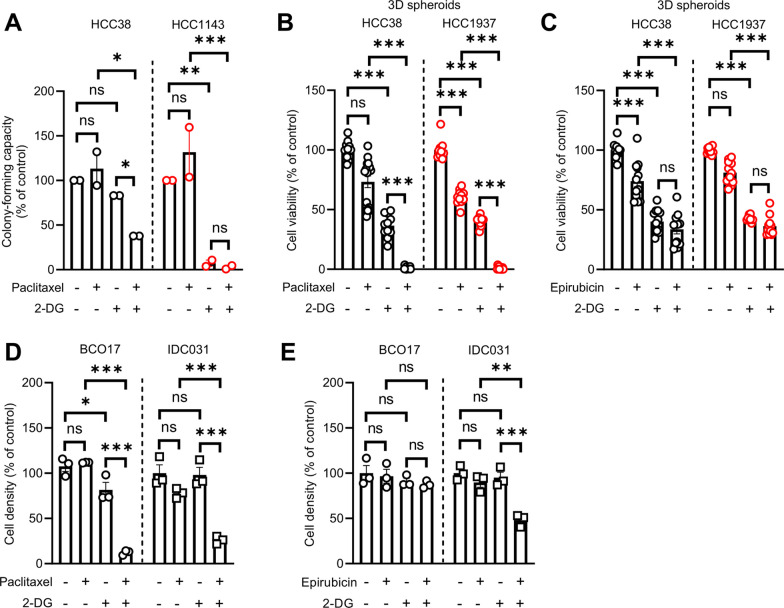
Methods: Human TNBC cell lines and patient-derived organoids were used in combination with real-time metabolic assays to evaluate the effect of NAC (paclitaxel and epirubicin) on tumor cell metabolism, in particular glycolysis. Diagnostic biopsies (pre-NAC) from patients with early TNBC were analyzed by bulk RNA-sequencing to evaluate the predictive value of a glycolysis-related gene signature.
Results: Paclitaxel induced a consistent metabolic switch to glycolysis, correlated with a reduced mitochondrial oxidative metabolism, in TNBC cells. In pre-NAC diagnostic biopsies from TNBC patients, glycolysis was found to be upregulated in non-responders. Furthermore, glycolysis inhibition greatly improved response to NAC in TNBC organoid models.
Conclusions: Our study pinpoints a metabolic adaptation to glycolysis as a mechanism driving resistance to NAC in TNBC. Our data pave the way for the use of glycolysis-related genes as predictive biomarkers for NAC response, as well as the development of inhibitors to overcome this glycolysis-driven resistance to NAC in human TNBC patients.
Keywords: Early triple negative breast cancer; Glycolysis; Metabolism; Neoadjuvant chemotherapy; Organoids; Therapy resistance.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Yau C, Osdoit M, van der Noordaa M, Shad S, Wei J, de Croze D, et al. Residual cancer burden after neoadjuvant chemotherapy and long-term survival outcomes in breast cancer: A multicentre pooled analysis of 5161 patients. Lancet Oncol. 2022;23(1):149–160. doi: 10.1016/S1470-2045(21)00589-1. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
