

Diverse cytomegalovirus US11 antagonism and MHC-A evasion strategies reveal a tit-for-tat coevolutionary arms race in hominids
- PMID: 38377192
- PMCID: PMC10907249
- DOI: 10.1073/pnas.2315985121
Diverse cytomegalovirus US11 antagonism and MHC-A evasion strategies reveal a tit-for-tat coevolutionary arms race in hominids
Abstract
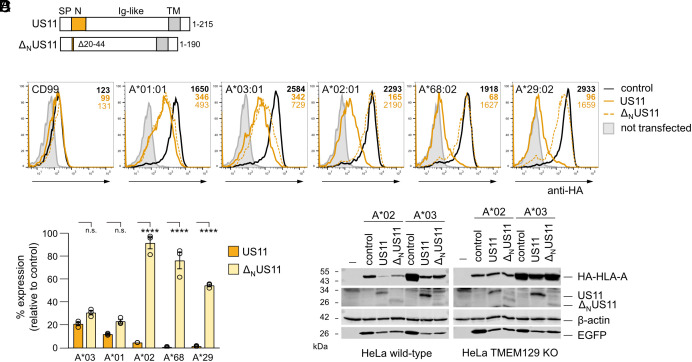
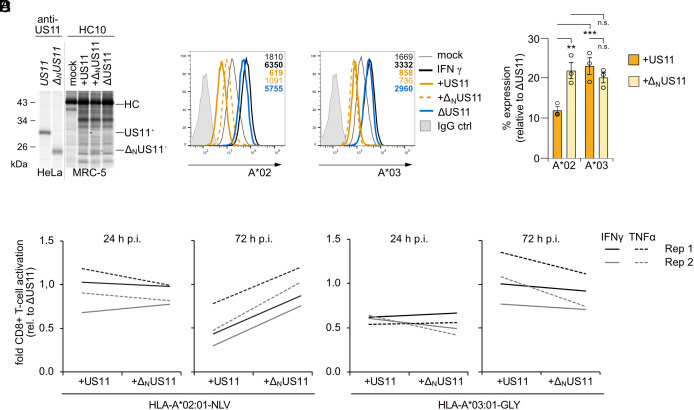
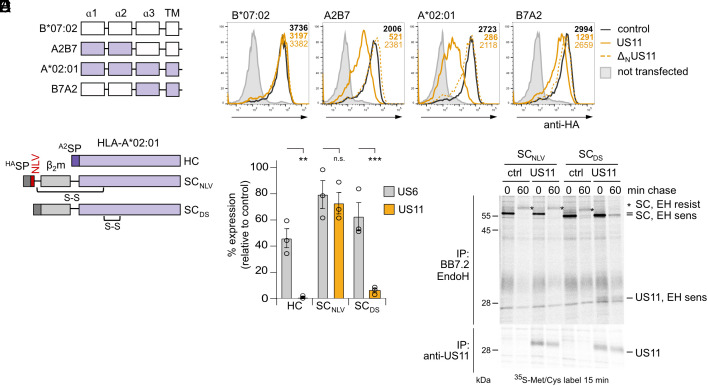
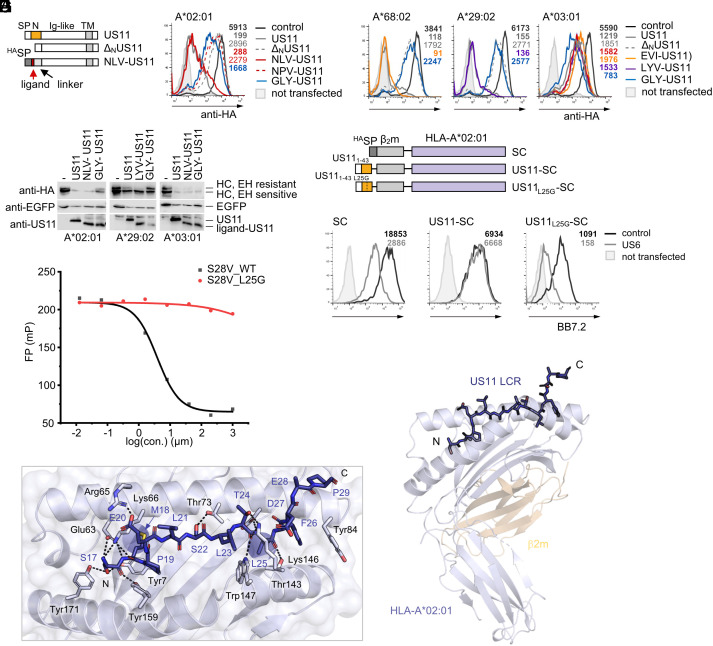
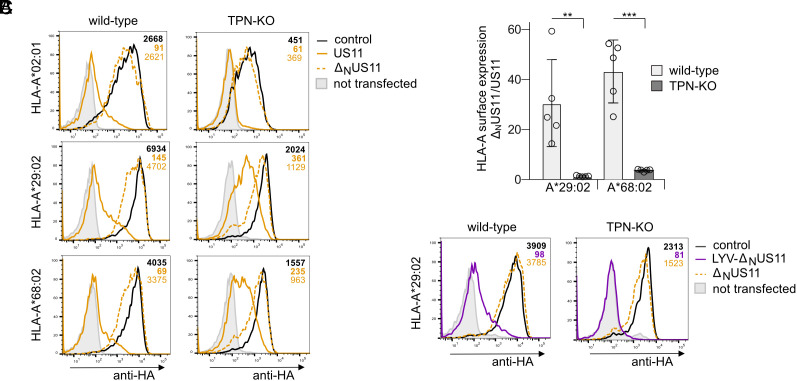
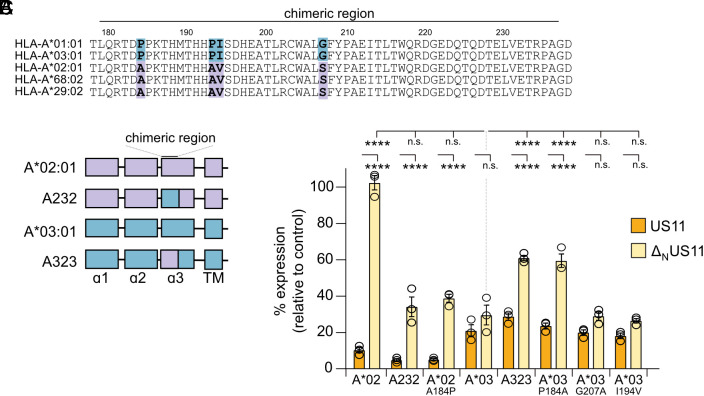
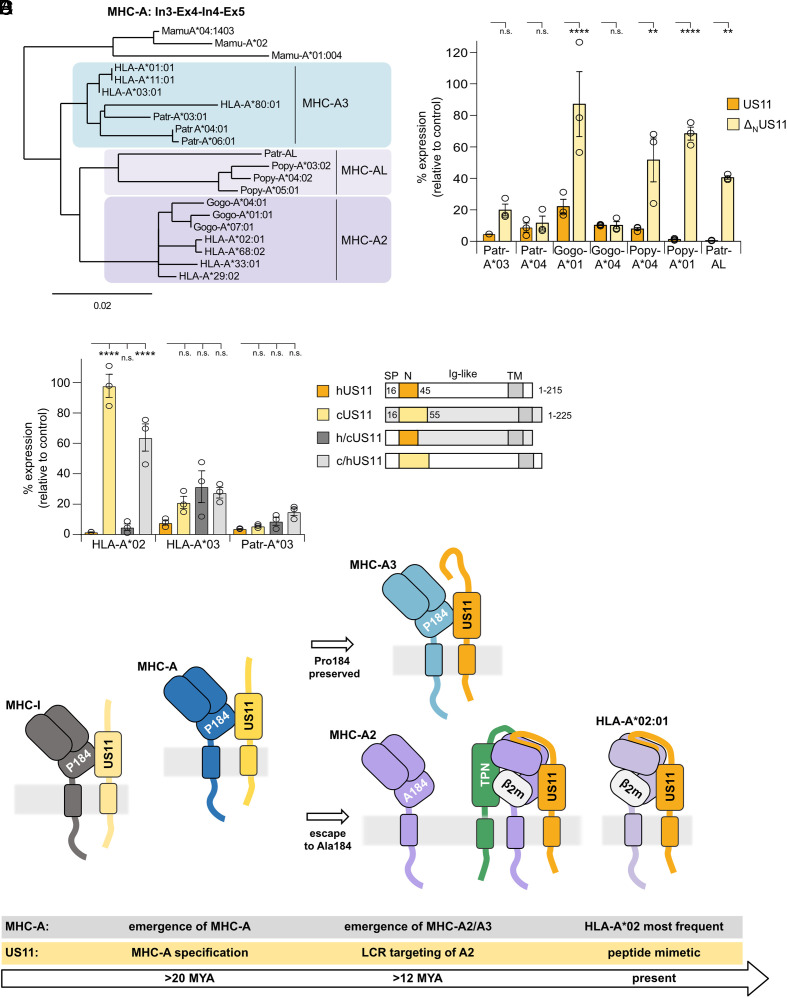
Recurrent, ancient arms races between viruses and hosts have shaped both host immunological defense strategies as well as viral countermeasures. One such battle is waged by the glycoprotein US11 encoded by the persisting human cytomegalovirus. US11 mediates degradation of major histocompatibility class I (MHC-I) molecules to prevent CD8+ T-cell activation. Here, we studied the consequences of the arms race between US11 and primate MHC-A proteins, leading us to uncover a tit-for-tat coevolution and its impact on MHC-A diversification. We found that US11 spurred MHC-A adaptation to evade viral antagonism: In an ancestor of great apes, the MHC-A A2 lineage acquired a Pro184Ala mutation, which confers resistance against the ancestral US11 targeting strategy. In response, US11 deployed a unique low-complexity region (LCR), which exploits the MHC-I peptide loading complex to target the MHC-A2 peptide-binding groove. In addition, the global spread of the human HLA-A*02 allelic family prompted US11 to employ a superior LCR strategy with an optimally fitting peptide mimetic that specifically antagonizes HLA-A*02. Thus, despite cytomegaloviruses low pathogenic potential, the increasing commitment of US11 to MHC-A has significantly promoted diversification of MHC-A in hominids.
Keywords: HLA-A; MHC class I; coevolution; cytomegalovirus; tapasin.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures
Similar articles
-
Multimodal HLA-I genotype regulation by human cytomegalovirus US10 and resulting surface patterning.Elife. 2024 Jun 20;13:e85560. doi: 10.7554/eLife.85560. Elife. 2024. PMID: 38900146 Free PMC article.
-
HLA-B locus products resist degradation by the human cytomegalovirus immunoevasin US11.PLoS Pathog. 2019 Sep 17;15(9):e1008040. doi: 10.1371/journal.ppat.1008040. eCollection 2019 Sep. PLoS Pathog. 2019. PMID: 31527904 Free PMC article.
-
Amino acid composition of alpha1/alpha2 domains and cytoplasmic tail of MHC class I molecules determine their susceptibility to human cytomegalovirus US11-mediated down-regulation.Eur J Immunol. 2003 Jun;33(6):1707-16. doi: 10.1002/eji.200323912. Eur J Immunol. 2003. PMID: 12778489
-
The HCMV gene products US2 and US11 target MHC class I molecules for degradation in the cytosol.Curr Top Microbiol Immunol. 2002;269:37-55. doi: 10.1007/978-3-642-59421-2_3. Curr Top Microbiol Immunol. 2002. PMID: 12224515 Review.
-
Inhibition of the MHC class II antigen presentation pathway by human cytomegalovirus.Curr Top Microbiol Immunol. 2002;269:101-15. doi: 10.1007/978-3-642-59421-2_7. Curr Top Microbiol Immunol. 2002. PMID: 12224504 Review.
Cited by
-
Multimodal HLA-I genotype regulation by human cytomegalovirus US10 and resulting surface patterning.Elife. 2024 Jun 20;13:e85560. doi: 10.7554/eLife.85560. Elife. 2024. PMID: 38900146 Free PMC article.
-
Human Cytomegalovirus Antigen Presentation by HLA-G in Infected Cells.HLA. 2025 May;105(5):e70089. doi: 10.1111/tan.70089. HLA. 2025. PMID: 40347012 Free PMC article.
-
Peptide-specific natural killer cell receptors.Oxf Open Immunol. 2025 Apr 17;6(1):iqaf003. doi: 10.1093/oxfimm/iqaf003. eCollection 2025. Oxf Open Immunol. 2025. PMID: 40297637 Free PMC article. Review.
References
-
- Berry R., Watson G. M., Jonjic S., Degli-Esposti M. A., Rossjohn J., Modulation of innate and adaptive immunity by cytomegaloviruses. Nat. Rev. Immunol. 20, 113–127 (2020). - PubMed
-
- Guma M., et al. , Imprint of human cytomegalovirus infection on the NK cell receptor repertoire. Blood 104, 3664–3671 (2004). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
