

The global RNA-RNA interactome of Klebsiella pneumoniae unveils a small RNA regulator of cell division
- PMID: 38377209
- PMCID: PMC10907235
- DOI: 10.1073/pnas.2317322121
The global RNA-RNA interactome of Klebsiella pneumoniae unveils a small RNA regulator of cell division
Abstract
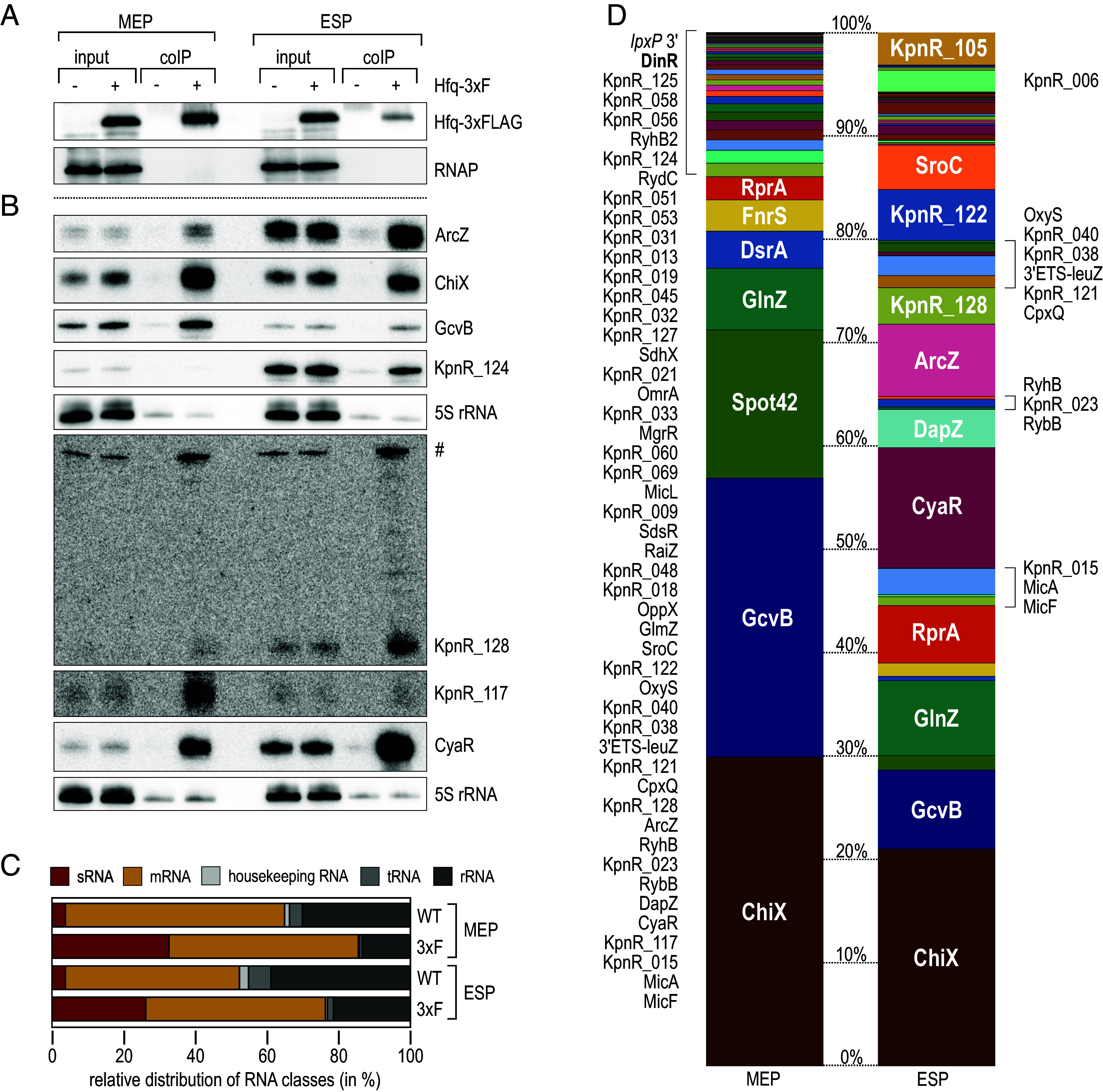
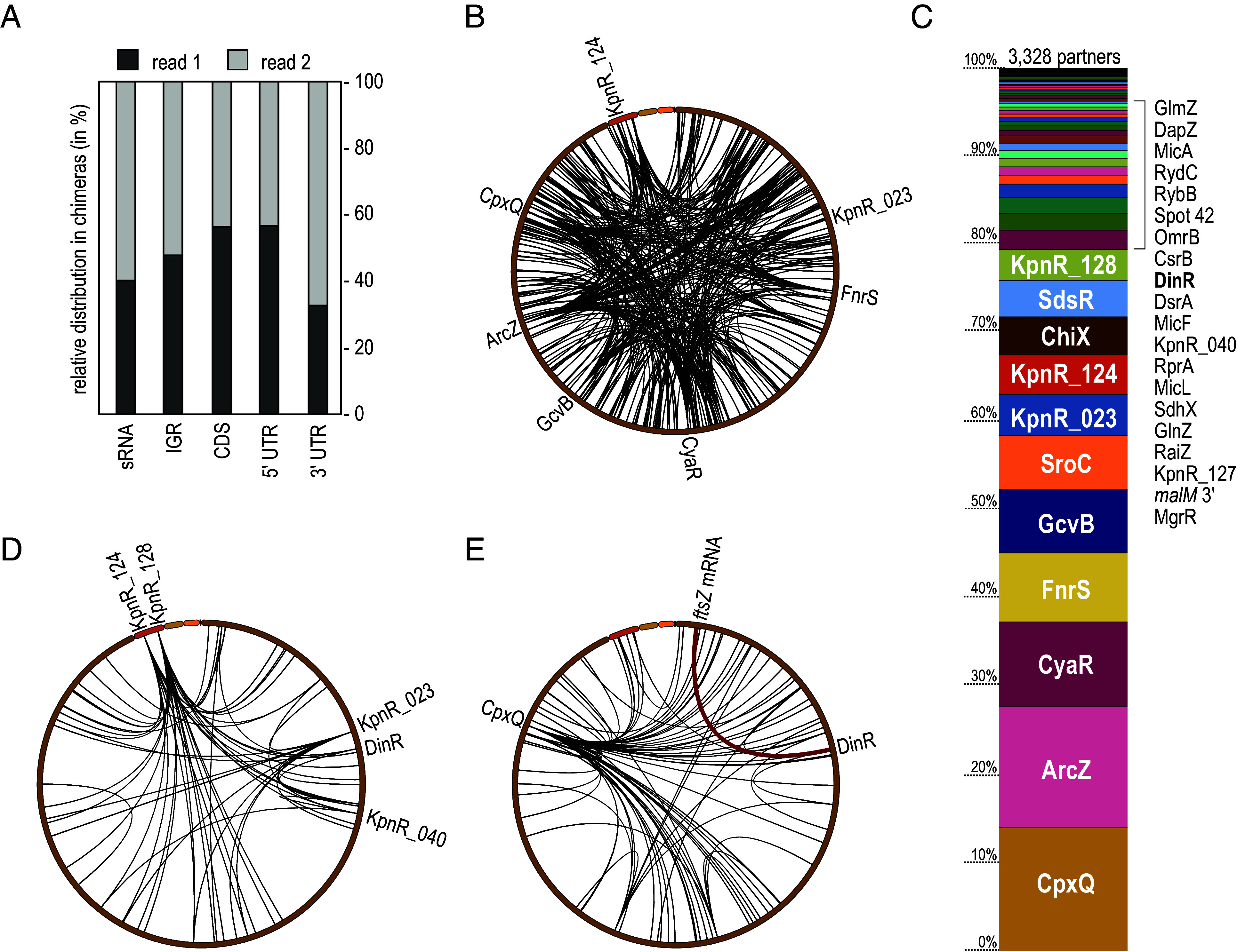
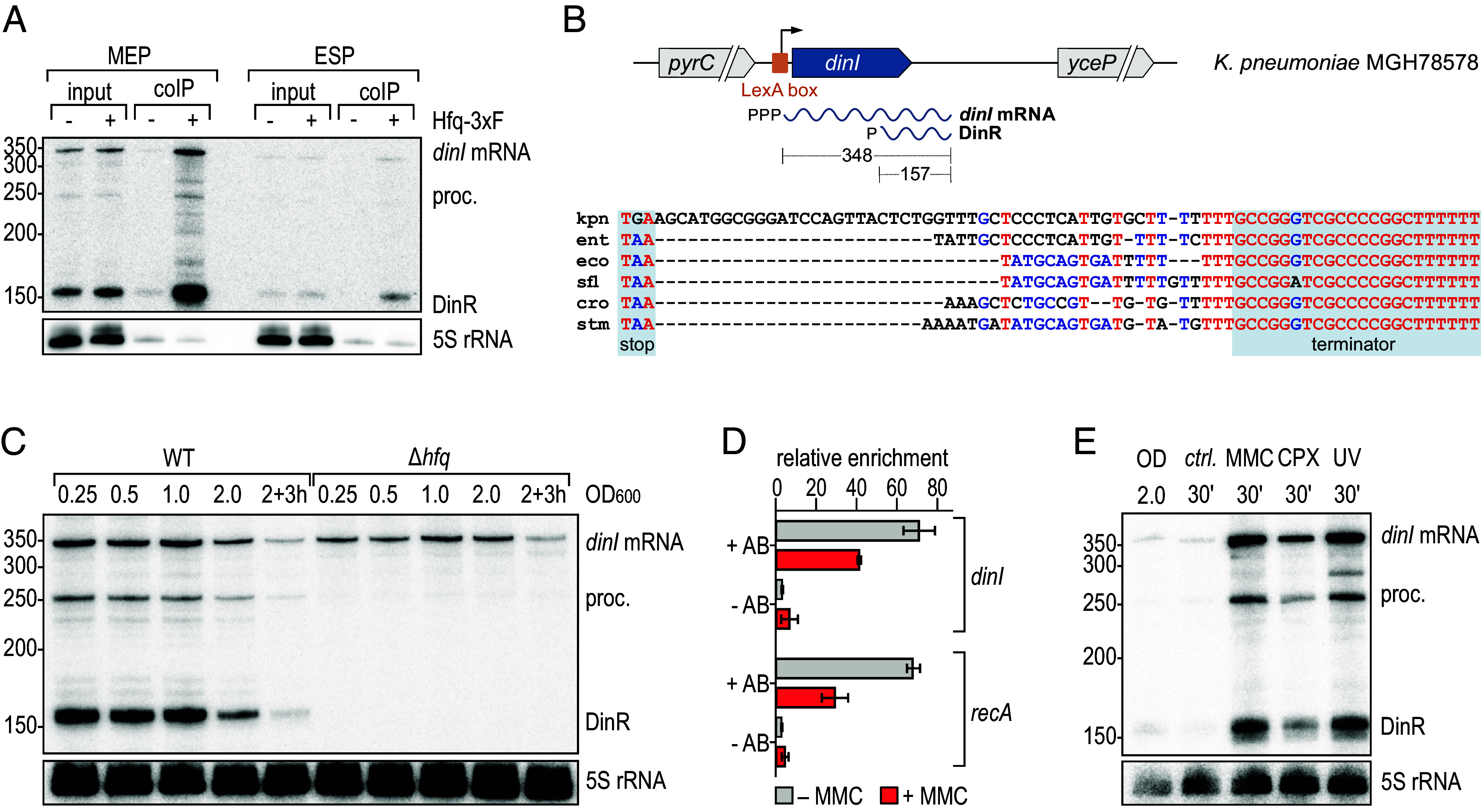
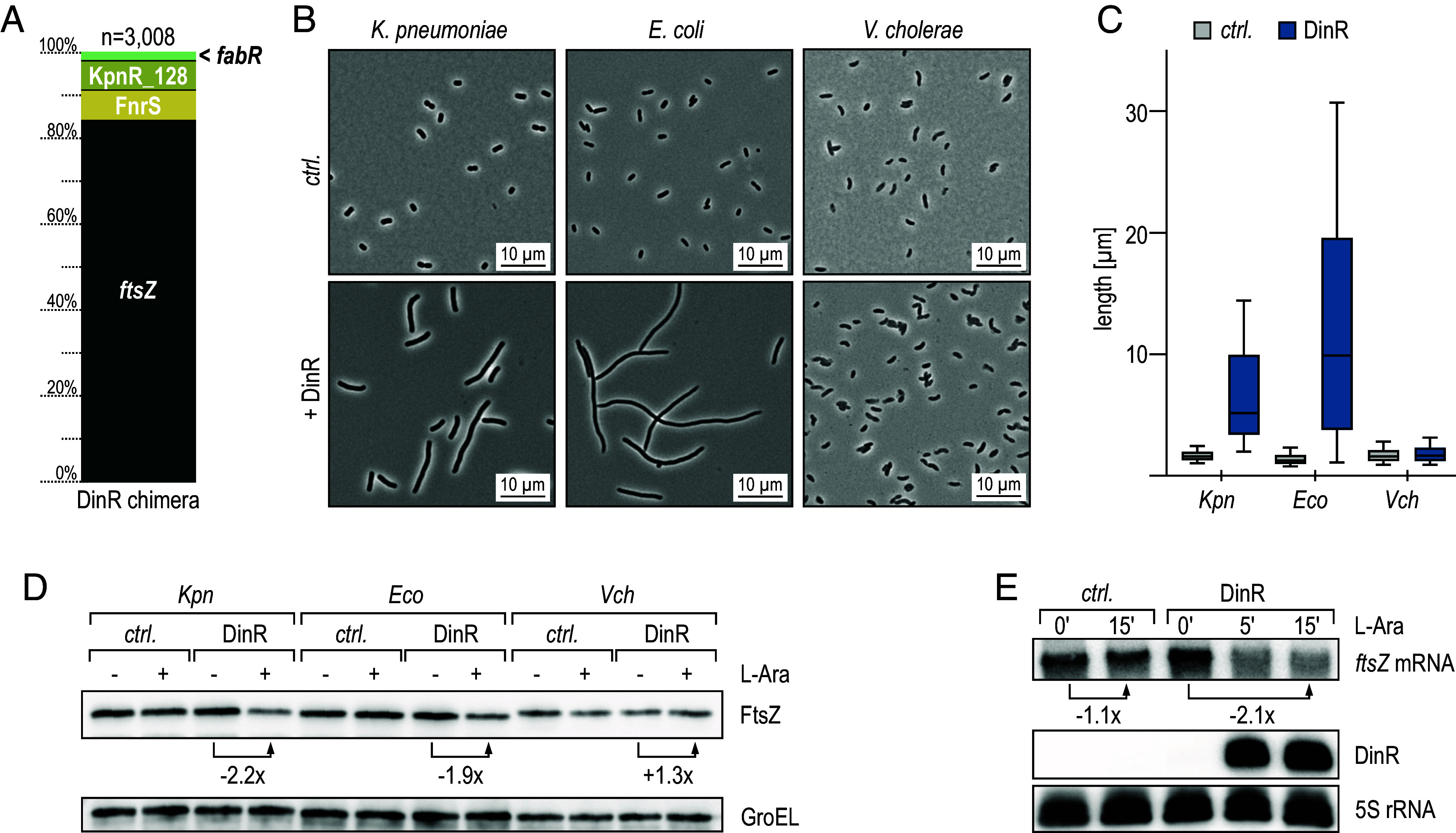
The ubiquitous RNA chaperone Hfq is involved in the regulation of key biological processes in many species across the bacterial kingdom. In the opportunistic human pathogen Klebsiella pneumoniae, deletion of the hfq gene affects the global transcriptome, virulence, and stress resistance; however, the ligands of the major RNA-binding protein in this species have remained elusive. In this study, we have combined transcriptomic, co-immunoprecipitation, and global RNA interactome analyses to compile an inventory of conserved and species-specific RNAs bound by Hfq and to monitor Hfq-mediated RNA-RNA interactions. In addition to dozens of RNA-RNA pairs, our study revealed an Hfq-dependent small regulatory RNA (sRNA), DinR, which is processed from the 3' terminal portion of dinI mRNA. Transcription of dinI is controlled by the master regulator of the SOS response, LexA. As DinR accumulates in K. pneumoniae in response to DNA damage, the sRNA represses translation of the ftsZ transcript by occupation of the ribosome binding site. Ectopic overexpression of DinR causes depletion of ftsZ mRNA and inhibition of cell division, while deletion of dinR antagonizes cell elongation in the presence of DNA damage. Collectively, our work highlights the important role of RNA-based gene regulation in K. pneumoniae and uncovers the central role of DinR in LexA-controlled division inhibition during the SOS response.
Keywords: Hfq; Klebsiella pneumoniae; RIL-seq; SOS response; small RNA.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures
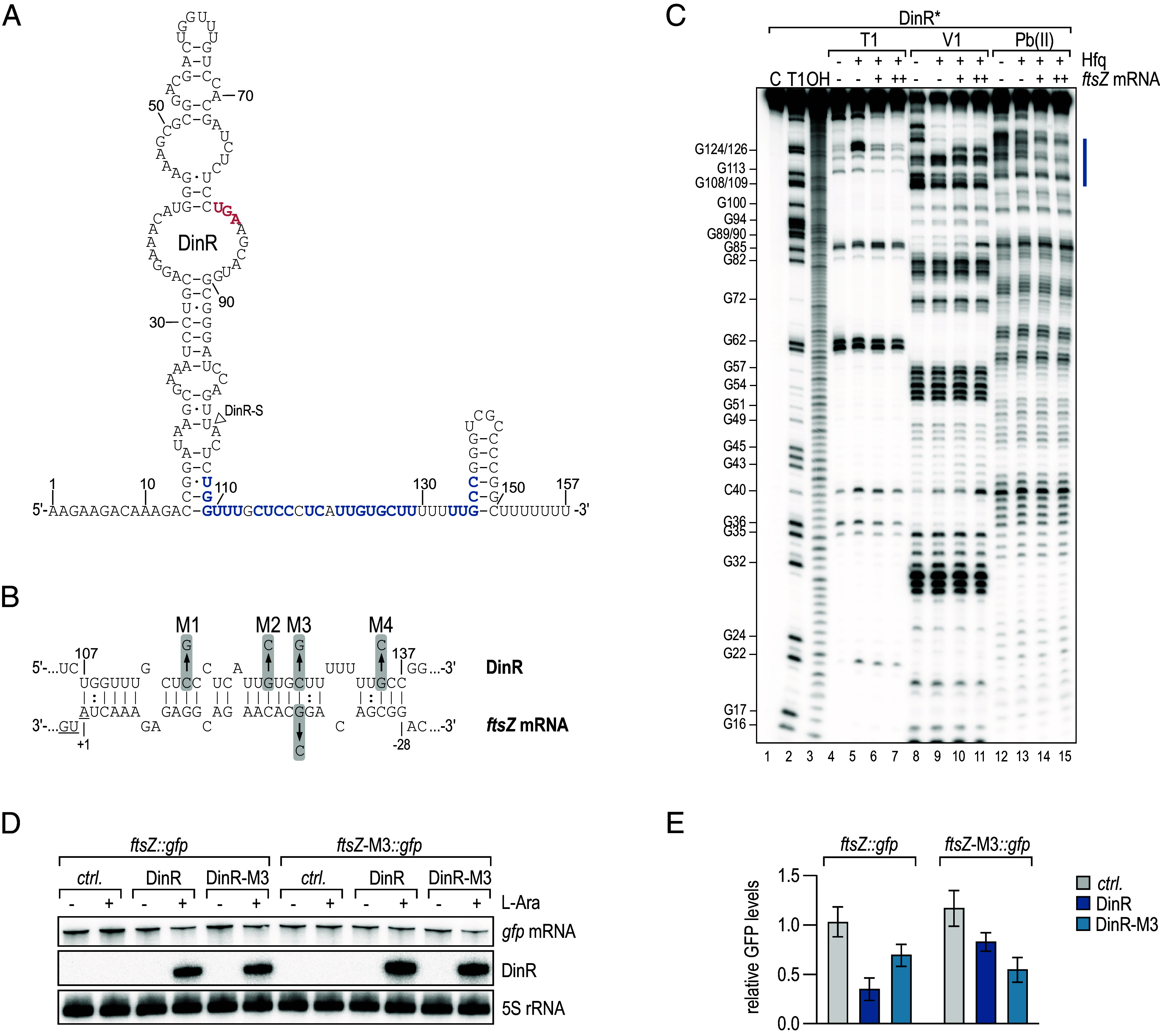
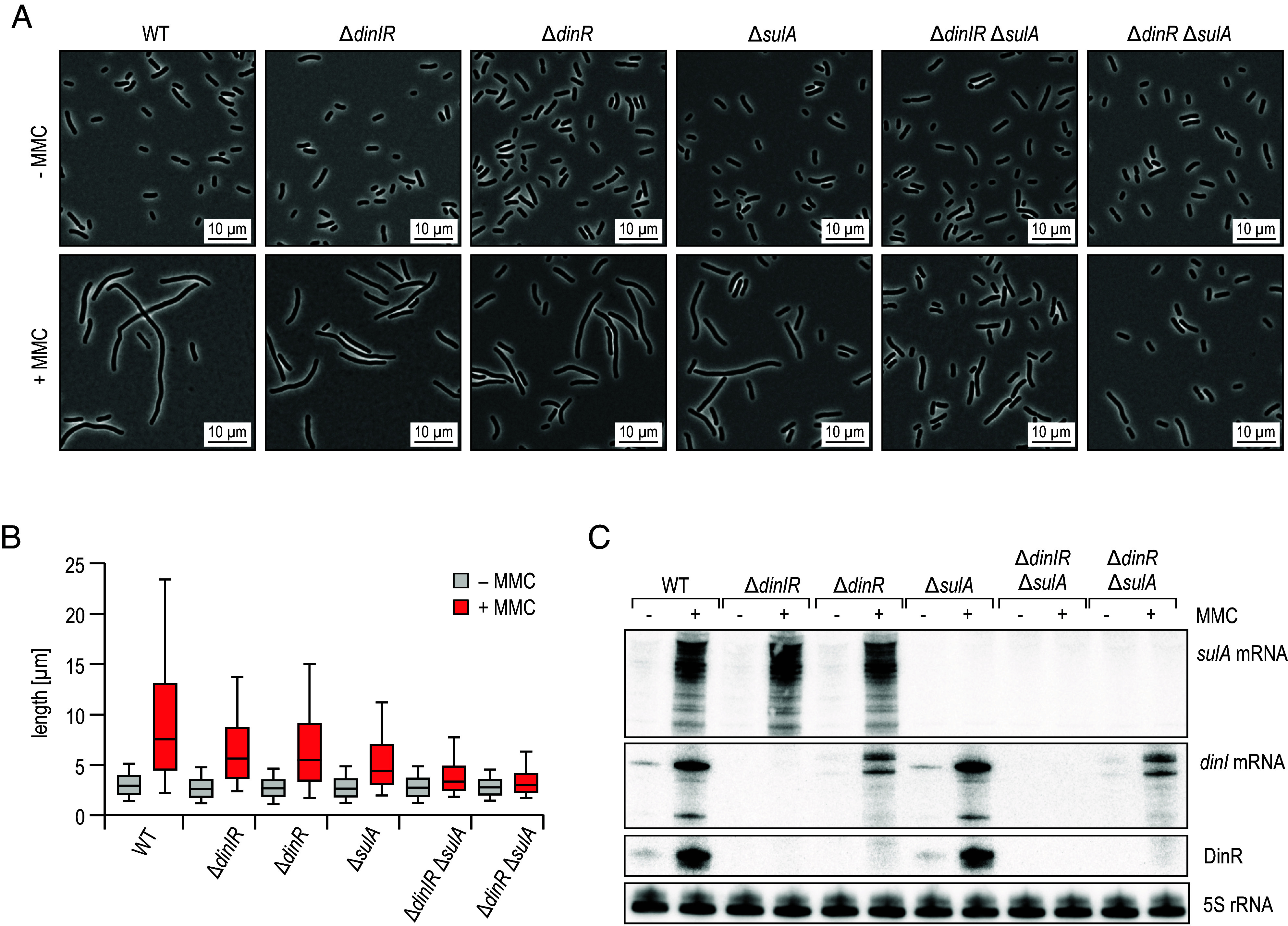
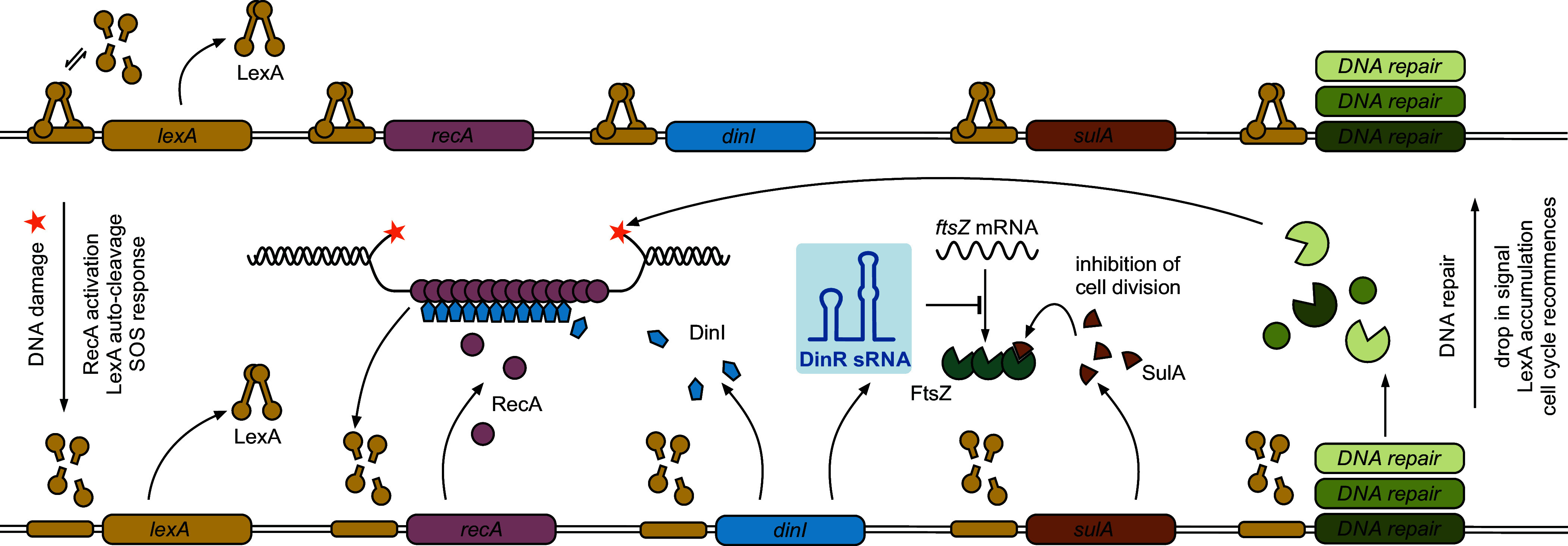
References
-
- Tacconelli E., et al. , Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 18, 318–327 (2018). - PubMed
-
- Sommer M. O. A., Munck C., Toft-Kehler R. V., Andersson D. I., Prediction of antibiotic resistance: Time for a new preclinical paradigm? Nat. Rev. Microbiol. 15, 689–696 (2017). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
