

Behavioral and Neurophysiological Implications of Pathological Human Tau Expression in Serotonin Neurons
- PMID: 38377680
- PMCID: PMC10921395
- DOI: 10.1021/acschemneuro.3c00626
Behavioral and Neurophysiological Implications of Pathological Human Tau Expression in Serotonin Neurons
Abstract
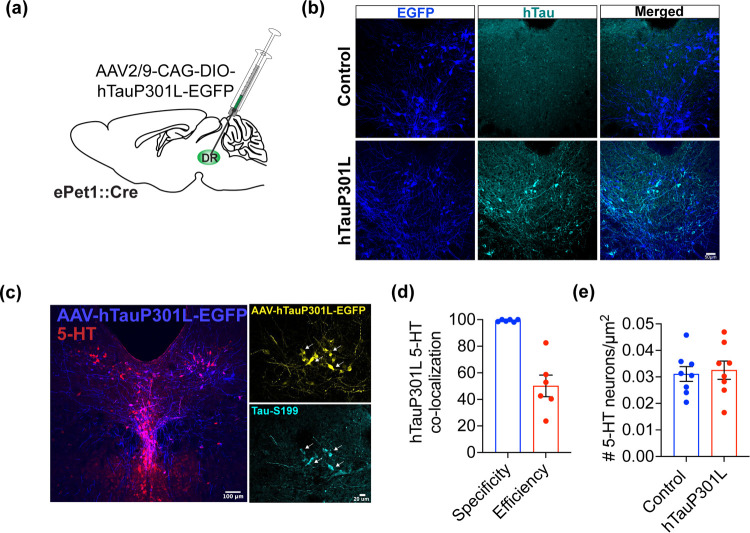
Alzheimer's disease (AD) is a progressive degenerative disorder that results in a severe loss of brain cells and irreversible cognitive decline. Memory problems are the most recognized symptoms of AD. However, approximately 90% of patients diagnosed with AD suffer from behavioral symptoms, including mood changes and social impairment years before cognitive dysfunction. Recent evidence indicates that the dorsal raphe nucleus (DRN) is among the initial regions that show tau pathology, which is a hallmark feature of AD. The DRN harbors serotonin (5-HT) neurons, which are critically involved in mood, social, and cognitive regulation. Serotonergic impairment early in the disease process may contribute to behavioral symptoms in AD. However, the mechanisms underlying vulnerability and contribution of the 5-HT system to AD progression remain unknown. Here, we performed behavioral and electrophysiological characterizations in mice expressing a phosphorylation-prone form of human tau (hTauP301L) in 5-HT neurons. We found that pathological tau expression in 5-HT neurons induces anxiety-like behavior and alterations in stress-coping strategies in female and male mice. Female mice also exhibited social disinhibition and mild cognitive impairment in response to 5-HT neuron-specific hTauP301L expression. Behavioral alterations were accompanied by disrupted 5-HT neuron physiology in female and male hTauP301L expressing mice with exacerbated excitability disruption in females only. These data provide mechanistic insights into the brain systems and symptoms impaired early in AD progression, which is critical for disease intervention.
Keywords: Alzheimer’s disease; anxiety; dorsal raphe nucleus; neuropsychiatric symptoms; serotonin (5-HT); tau.
Conflict of interest statement
The authors declare no competing financial interest.
Figures




Similar articles
-
Human tau-overexpressing mice recapitulate brainstem involvement and neuropsychiatric features of early Alzheimer's disease.Acta Neuropathol Commun. 2023 Apr 3;11(1):57. doi: 10.1186/s40478-023-01546-5. Acta Neuropathol Commun. 2023. PMID: 37009893 Free PMC article.
-
Dorsal raphe nucleus-hippocampus serotonergic circuit underlies the depressive and cognitive impairments in 5×FAD male mice.Transl Neurodegener. 2024 Jul 24;13(1):34. doi: 10.1186/s40035-024-00425-w. Transl Neurodegener. 2024. PMID: 39044270 Free PMC article.
-
Quantifying the accretion of hyperphosphorylated tau in the locus coeruleus and dorsal raphe nucleus: the pathological building blocks of early Alzheimer's disease.Neuropathol Appl Neurobiol. 2017 Aug;43(5):393-408. doi: 10.1111/nan.12387. Epub 2017 Mar 31. Neuropathol Appl Neurobiol. 2017. PMID: 28117917 Free PMC article.
-
Putative pathological mechanisms of late-life depression and Alzheimer's disease.Brain Res. 2023 Aug 15;1813:148423. doi: 10.1016/j.brainres.2023.148423. Epub 2023 May 25. Brain Res. 2023. PMID: 37244602 Review.
-
Ascending monoaminergic systems alterations in Alzheimer's disease. translating basic science into clinical care.Neurosci Biobehav Rev. 2013 Sep;37(8):1363-79. doi: 10.1016/j.neubiorev.2013.05.008. Epub 2013 May 24. Neurosci Biobehav Rev. 2013. PMID: 23707776 Review.
Cited by
-
Deciphering the Functions of Raphe-Hippocampal Serotonergic and Glutamatergic Circuits and Their Deficits in Alzheimer's Disease.Int J Mol Sci. 2025 Jan 30;26(3):1234. doi: 10.3390/ijms26031234. Int J Mol Sci. 2025. PMID: 39941002 Free PMC article. Review.
-
SSRIs reduce plasma tau and restore dorsal raphe metabolism in Alzheimer's disease.Alzheimers Dement. 2025 Feb;21(2):e14579. doi: 10.1002/alz.14579. Alzheimers Dement. 2025. PMID: 39935329 Free PMC article.
References
-
- Hyman B. T.; Phelps C. H.; Beach T. G.; Bigio E. H.; Cairns N. J.; Carrillo M. C.; Dickson D. W.; Duyckaerts C.; Frosch M. P.; Masliah E.; Mirra S. S.; Nelson P. T.; Schneider J. A.; Thal D. R.; Thies B.; Trojanowski J. Q.; Vinters H. V.; Montine T. J. National Institute on Aging-Alzheimer’s Association Guidelines for the Neuropathologic Assessment of Alzheimer’s Disease. Alzheimer’s Dementia 2012, 8 (1), 1–13. 10.1016/j.jalz.2011.10.007. - DOI - PMC - PubMed
-
- Ringman J. M.; Liang L.-J.; Zhou Y.; Vangala S.; Teng E.; Kremen S.; Wharton D.; Goate A.; Marcus D. S.; Farlow M.; Ghetti B.; McDade E.; Masters C. L.; Mayeux R. P.; Rossor M.; Salloway S.; Schofield P. R.; Cummings J. L.; Buckles V.; Bateman R.; Morris J. C.; Early Behavioural Changes in Familial Alzheimer’s Disease in the Dominantly Inherited Alzheimer Network. Brain 2015, 138 (Pt 4), 1036–1045. 10.1093/brain/awv004. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
