

IgG is an aging factor that drives adipose tissue fibrosis and metabolic decline
- PMID: 38378001
- PMCID: PMC11070064
- DOI: 10.1016/j.cmet.2024.01.015
IgG is an aging factor that drives adipose tissue fibrosis and metabolic decline
Abstract
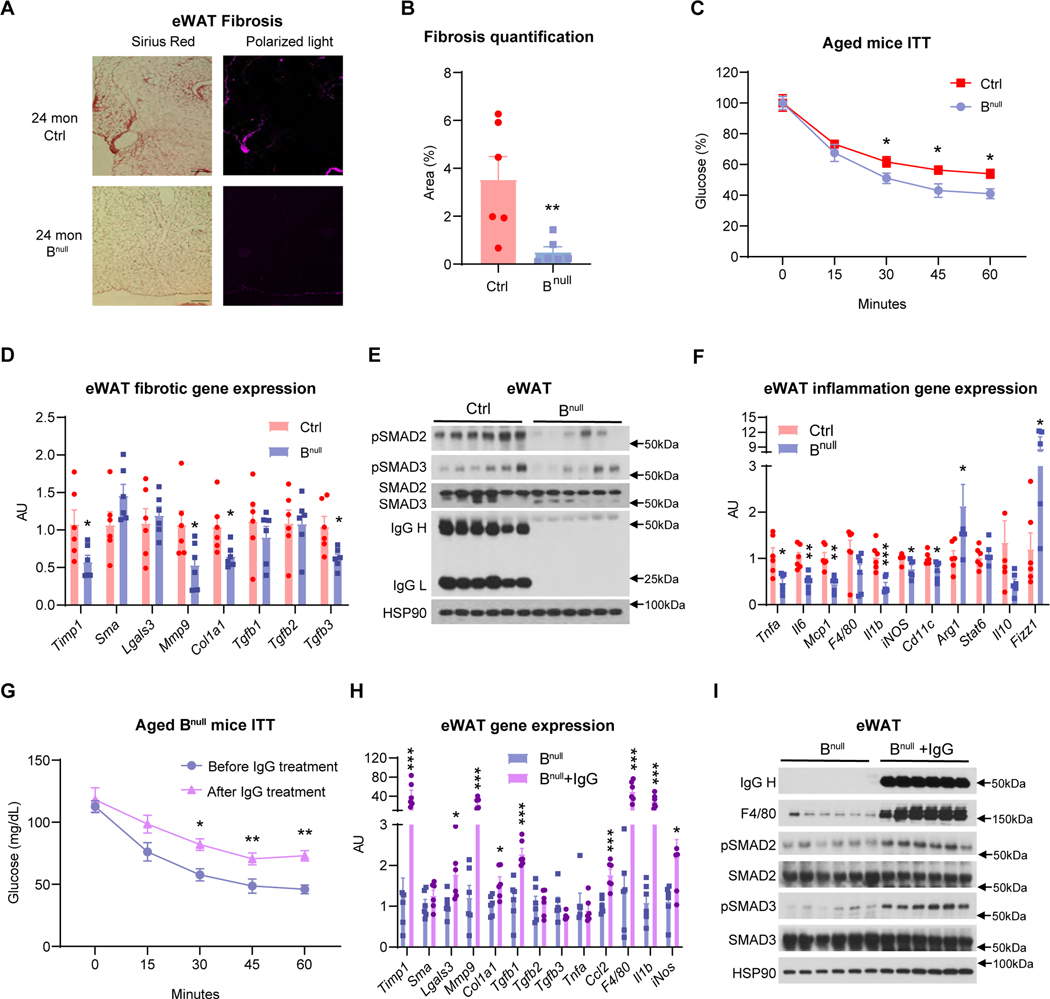
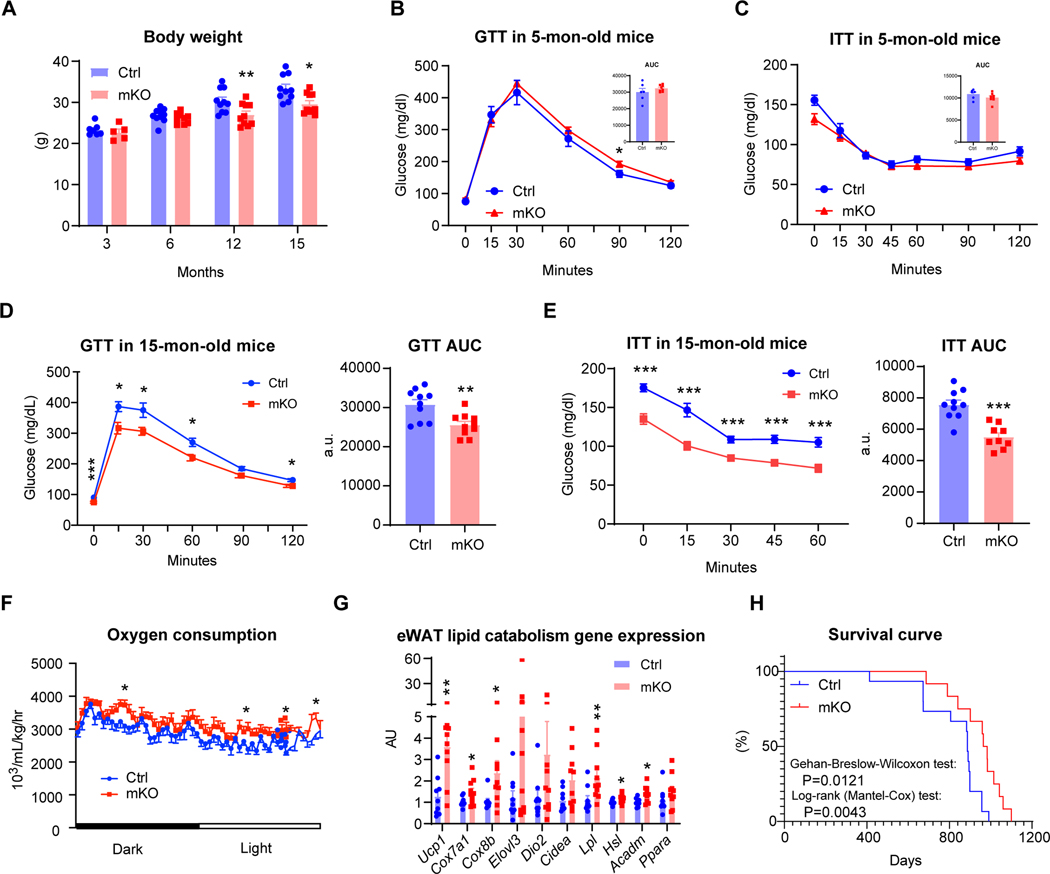
Aging is underpinned by pronounced metabolic decline; however, the drivers remain obscure. Here, we report that IgG accumulates during aging, particularly in white adipose tissue (WAT), to impair adipose tissue function and metabolic health. Caloric restriction (CR) decreases IgG accumulation in WAT, whereas replenishing IgG counteracts CR's metabolic benefits. IgG activates macrophages via Ras signaling and consequently induces fibrosis in WAT through the TGF-β/SMAD pathway. Consistently, B cell null mice are protected from aging-associated WAT fibrosis, inflammation, and insulin resistance, unless exposed to IgG. Conditional ablation of the IgG recycling receptor, neonatal Fc receptor (FcRn), in macrophages prevents IgG accumulation in aging, resulting in prolonged healthspan and lifespan. Further, targeting FcRn by antisense oligonucleotide restores WAT integrity and metabolic health in aged mice. These findings pinpoint IgG as a hidden culprit in aging and enlighten a novel strategy to rejuvenate metabolic health.
Keywords: IgG; adipose tissue; aging; fibrosis; metabolic dysfunction.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A patent application is pending by Columbia University.
Figures





References
-
- Hildrum B, Mykletun A, Hole T, Midthjell K, and Dahl AA. (2007). Age-specific prevalence of the metabolic syndrome defined by the International Diabetes Federation and the National Cholesterol Education Program: the Norwegian HUNT 2 study. BMC public health 7, 220. 10.1186/1471-2458-7-220. - DOI - PMC - PubMed
-
- Grover SA, Kaouache M, Rempel P, Joseph L, Dawes M, Lau DC, and Lowensteyn I. (2015). Years of life lost and healthy life-years lost from diabetes and cardiovascular disease in overweight and obese people: a modelling study. Lancet Diabetes Endocrinol 3, 114–122. 10.1016/S2213-8587(14)70229-3. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
