

Differential leaf flooding resilience in Arabidopsis thaliana is controlled by ethylene signaling-activated and age-dependent phosphorylation of ORESARA1
- PMID: 38379284
- PMCID: PMC11211547
- DOI: 10.1016/j.xplc.2024.100848
Differential leaf flooding resilience in Arabidopsis thaliana is controlled by ethylene signaling-activated and age-dependent phosphorylation of ORESARA1
Abstract
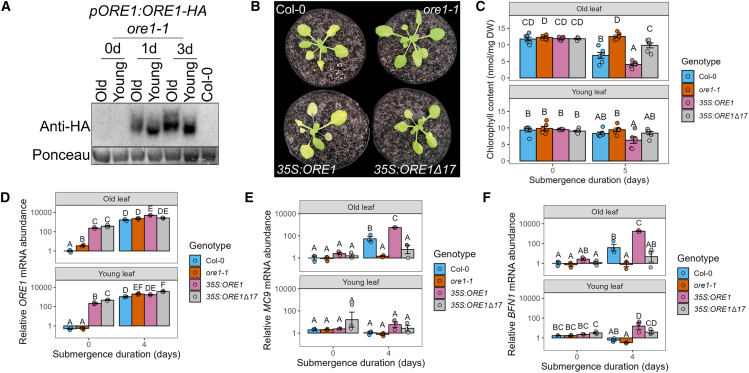
The phytohormone ethylene is a major regulator of plant adaptive responses to flooding. In flooded plant tissues, ethylene quickly increases to high concentrations owing to its low solubility and diffusion rates in water. Ethylene accumulation in submerged plant tissues makes it a reliable cue for triggering flood acclimation responses, including metabolic adjustments to cope with flood-induced hypoxia. However, persistent ethylene accumulation also accelerates leaf senescence. Stress-induced senescence hampers photosynthetic capacity and stress recovery. In submerged Arabidopsis, senescence follows a strict age-dependent pattern starting with the older leaves. Although mechanisms underlying ethylene-mediated senescence have been uncovered, it is unclear how submerged plants avoid indiscriminate breakdown of leaves despite high systemic ethylene accumulation. We demonstrate that although submergence triggers leaf-age-independent activation of ethylene signaling via EIN3 in Arabidopsis, senescence is initiated only in old leaves. EIN3 stabilization also leads to overall transcript and protein accumulation of the senescence-promoting transcription factor ORESARA1 (ORE1) in both old and young leaves during submergence. However, leaf-age-dependent senescence can be explained by ORE1 protein activation via phosphorylation specifically in old leaves, independent of the previously identified age-dependent control of ORE1 via miR164. A systematic analysis of the roles of the major flooding stress cues and signaling pathways shows that only the combination of ethylene and darkness is sufficient to mimic submergence-induced senescence involving ORE1 accumulation and phosphorylation. Hypoxia, most often associated with flooding stress in plants, appears to have no role in these processes. Our results reveal a mechanism by which plants regulate the speed and pattern of senescence during environmental stresses such as flooding. Age-dependent ORE1 activity ensures that older, expendable leaves are dismantled first, thus prolonging the life of younger leaves and meristematic tissues that are vital to whole-plant survival.
Keywords: abiotic stress; ethylene; flooding; hypoxia; senescence.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures






References
-
- Alonso J.M., Hirayama T., Roman G., Nourizadeh S., Ecker J.R. EIN2, a Bifunctional Transducer of Ethylene and Stress Responses in Arabidopsis. Science. 1999;284:2148–2152. - PubMed
-
- Alpuerto J.B., Hussain R.M.F., Fukao T. The key regulator of submergence tolerance, SUB1A, promotes photosynthetic and metabolic recovery from submergence damage in rice leaves. Plant Cell Environ. 2016;39:672–684. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
