

Regional centromere configuration in the fungal pathogens of the Pneumocystis genus
- PMID: 38380929
- PMCID: PMC10936427
- DOI: 10.1128/mbio.03185-23
Regional centromere configuration in the fungal pathogens of the Pneumocystis genus
Abstract
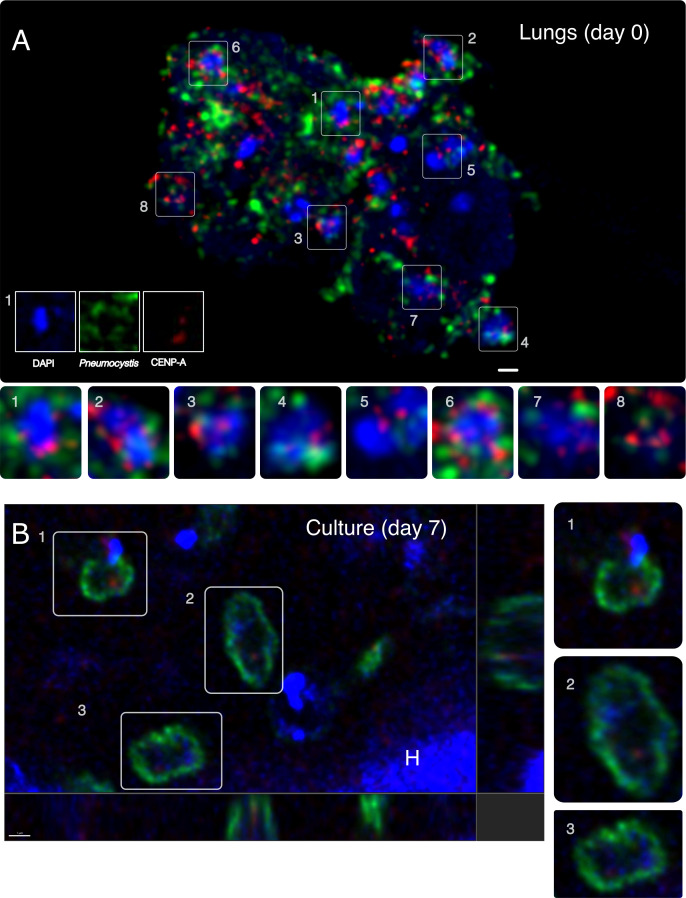
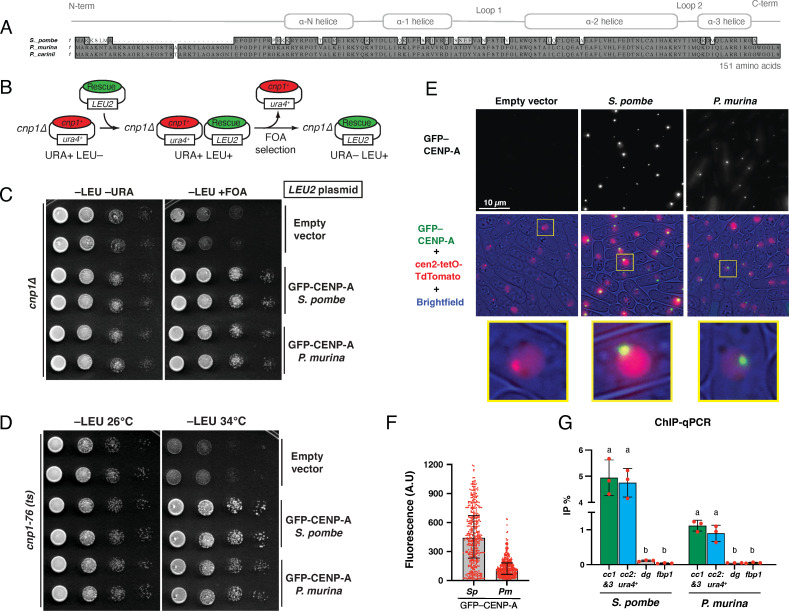
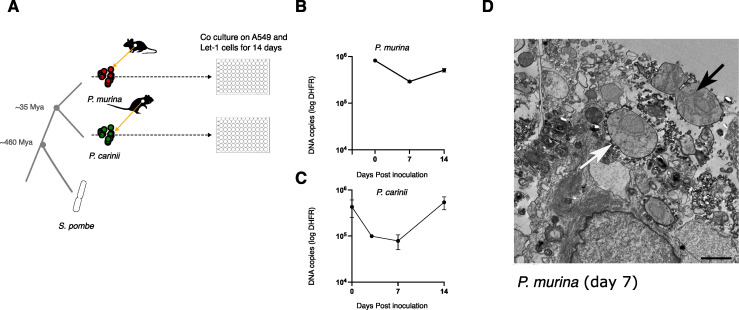
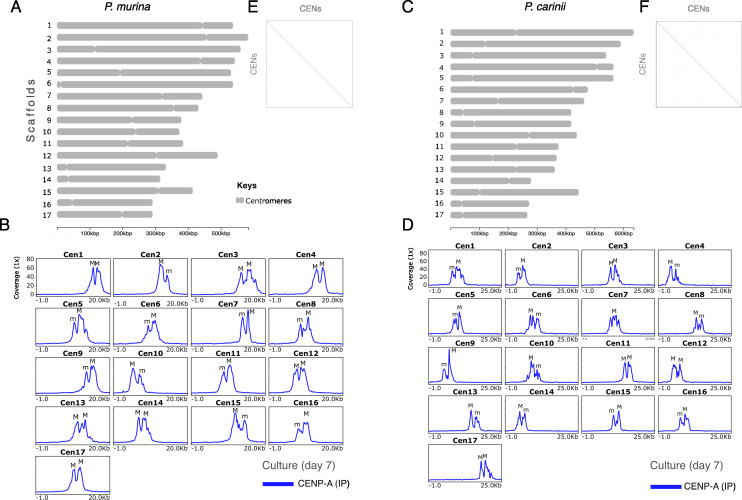
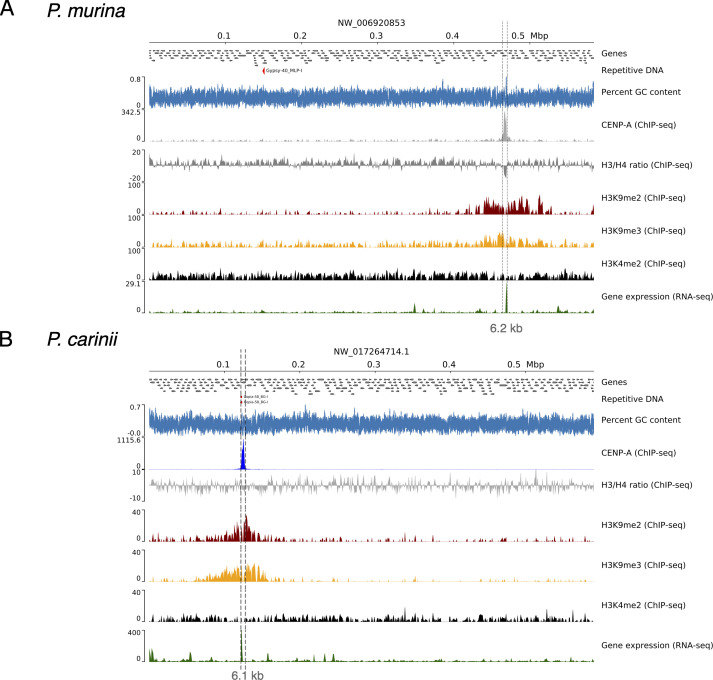
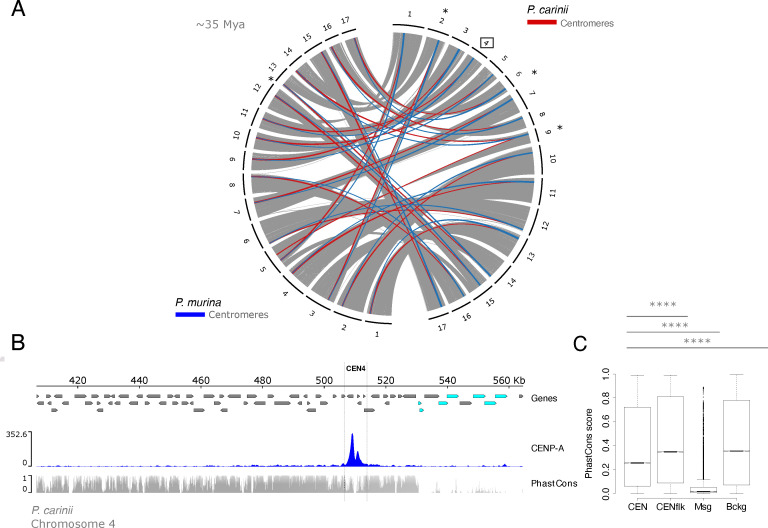
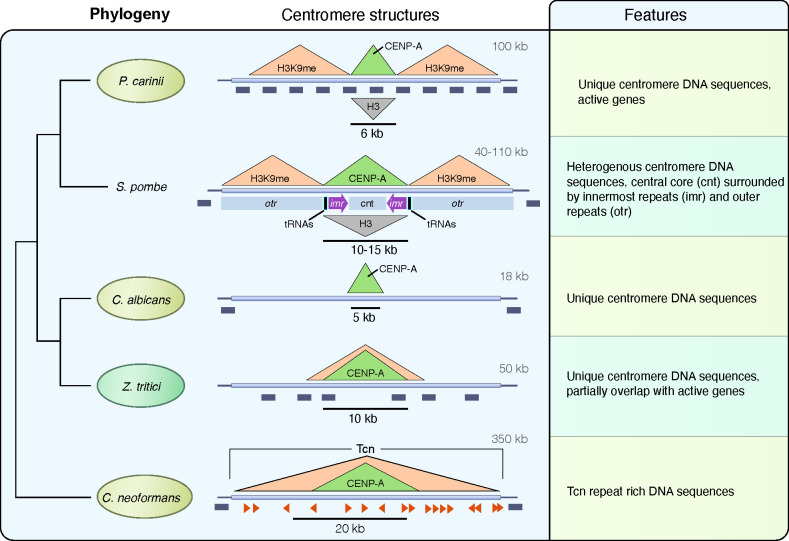
Centromeres are constricted chromosomal regions that are essential for cell division. In eukaryotes, centromeres display a remarkable architectural and genetic diversity. The basis of centromere-accelerated evolution remains elusive. Here, we focused on Pneumocystis species, a group of mammalian-specific fungal pathogens that form a sister taxon with that of the Schizosaccharomyces pombe, an important genetic model for centromere biology research. Methods allowing reliable continuous culture of Pneumocystis species do not currently exist, precluding genetic manipulation. CENP-A, a variant of histone H3, is the epigenetic marker that defines centromeres in most eukaryotes. Using heterologous complementation, we show that the Pneumocystis CENP-A ortholog is functionally equivalent to CENP-ACnp1 of S. pombe. Using organisms from a short-term in vitro culture or infected animal models and chromatin immunoprecipitation (ChIP)-Seq, we identified CENP-A bound regions in two Pneumocystis species that diverged ~35 million years ago. Each species has a unique short regional centromere (<10 kb) flanked by heterochromatin in 16-17 monocentric chromosomes. They span active genes and lack conserved DNA sequence motifs and repeats. These features suggest an epigenetic specification of centromere function. Analysis of centromeric DNA across multiple Pneumocystis species suggests a vertical transmission at least 100 million years ago. The common ancestry of Pneumocystis and S. pombe centromeres is untraceable at the DNA level, but the overall architectural similarity could be the result of functional constraint for successful chromosomal segregation.IMPORTANCEPneumocystis species offer a suitable genetic system to study centromere evolution in pathogens because of their phylogenetic proximity with the non-pathogenic yeast S. pombe, a popular model for cell biology. We used this system to explore how centromeres have evolved after the divergence of the two clades ~ 460 million years ago. To address this question, we established a protocol combining short-term culture and ChIP-Seq to characterize centromeres in multiple Pneumocystis species. We show that Pneumocystis have short epigenetic centromeres that function differently from those in S. pombe.
Keywords: chromosome segregation; evolution; genetics; genome organization; opportunistic fungi.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Update of
-
The Host Adapted Fungal Pathogens of Pneumocystis Genus Utilize Genic Regional Centromeres.bioRxiv [Preprint]. 2023 May 12:2023.05.12.540427. doi: 10.1101/2023.05.12.540427. bioRxiv. 2023. Update in: mBio. 2024 Mar 13;15(3):e0318523. doi: 10.1128/mbio.03185-23. PMID: 37425787 Free PMC article. Updated. Preprint.
Similar articles
-
The Host Adapted Fungal Pathogens of Pneumocystis Genus Utilize Genic Regional Centromeres.bioRxiv [Preprint]. 2023 May 12:2023.05.12.540427. doi: 10.1101/2023.05.12.540427. bioRxiv. 2023. Update in: mBio. 2024 Mar 13;15(3):e0318523. doi: 10.1128/mbio.03185-23. PMID: 37425787 Free PMC article. Updated. Preprint.
-
Heterochromatin and RNAi regulate centromeres by protecting CENP-A from ubiquitin-mediated degradation.PLoS Genet. 2018 Aug 8;14(8):e1007572. doi: 10.1371/journal.pgen.1007572. eCollection 2018 Aug. PLoS Genet. 2018. PMID: 30089114 Free PMC article.
-
Interspecies conservation of organisation and function between nonhomologous regional centromeres.Nat Commun. 2019 May 28;10(1):2343. doi: 10.1038/s41467-019-09824-4. Nat Commun. 2019. PMID: 31138803 Free PMC article.
-
"Lessons from the extremes: Epigenetic and genetic regulation in point monocentromere and holocentromere establishment on artificial chromosomes".Exp Cell Res. 2020 May 15;390(2):111974. doi: 10.1016/j.yexcr.2020.111974. Epub 2020 Mar 26. Exp Cell Res. 2020. PMID: 32222413 Review.
-
Two distinct pathways responsible for the loading of CENP-A to centromeres in the fission yeast cell cycle.Philos Trans R Soc Lond B Biol Sci. 2005 Mar 29;360(1455):595-606; discussion 606-7. doi: 10.1098/rstb.2004.1614. Philos Trans R Soc Lond B Biol Sci. 2005. PMID: 15897182 Free PMC article. Review.
Cited by
-
Extracellular Vesicle-Mediated Delivery of Genetic Material for Transformation and CRISPR/Cas9-based Gene Editing in Pneumocystis murina.bioRxiv [Preprint]. 2025 Jun 17:2025.06.17.660080. doi: 10.1101/2025.06.17.660080. bioRxiv. 2025. PMID: 40667333 Free PMC article. Preprint.
-
Long-read genomics reveal extensive nuclear-specific evolution and allele-specific expression in a dikaryotic fungus.Genome Res. 2025 Jun 2;35(6):1364-1376. doi: 10.1101/gr.280359.124. Genome Res. 2025. PMID: 40216555 Free PMC article.
References
-
- Sankaranarayanan SR, Ianiri G, Coelho MA, Reza MH, Thimmappa BC, Ganguly P, Vadnala RN, Sun S, Siddharthan R, Tellgren-Roth C, Dawson TL, Heitman J, Sanyal K. 2020. Loss of centromere function drives karyotype evolution in closely related Malassezia species. Elife 9:e53944. doi:10.7554/eLife.53944 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
