

Flipping the script: Advances in understanding how and why P4-ATPases flip lipid across membranes
- PMID: 38382846
- PMCID: PMC12424579
- DOI: 10.1016/j.bbamcr.2024.119700
Flipping the script: Advances in understanding how and why P4-ATPases flip lipid across membranes
Abstract
Type IV P-type ATPases (P4-ATPases) are a family of transmembrane enzymes that translocate lipid substrates from the outer to the inner leaflet of biological membranes and thus create an asymmetrical distribution of lipids within membranes. On the cellular level, this asymmetry is essential for maintaining the integrity and functionality of biological membranes, creating platforms for signaling events and facilitating vesicular trafficking. On the organismal level, this asymmetry has been shown to be important in maintaining blood homeostasis, liver metabolism, neural development, and the immune response. Indeed, dysregulation of P4-ATPases has been linked to several diseases; including anemia, cholestasis, neurological disease, and several cancers. This review will discuss the evolutionary transition of P4-ATPases from cation pumps to lipid flippases, the new lipid substrates that have been discovered, the significant advances that have been achieved in recent years regarding the structural mechanisms underlying the recognition and flipping of specific lipids across biological membranes, and the consequences of P4-ATPase dysfunction on cellular and physiological functions. Additionally, we emphasize the requirement for additional research to comprehensively understand the involvement of flippases in cellular physiology and disease and to explore their potential as targets for therapeutics in treating a variety of illnesses. The discussion in this review will primarily focus on the budding yeast, C. elegans, and mammalian P4-ATPases.
Keywords: Flippase; Glucosylceramide; Membrane asymmetry; P-type ATPase; Phosphatidylserine; Vesicular transport.
Copyright © 2024 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Todd R. Graham reports financial support was provided by the National Institutes of Health.
Figures


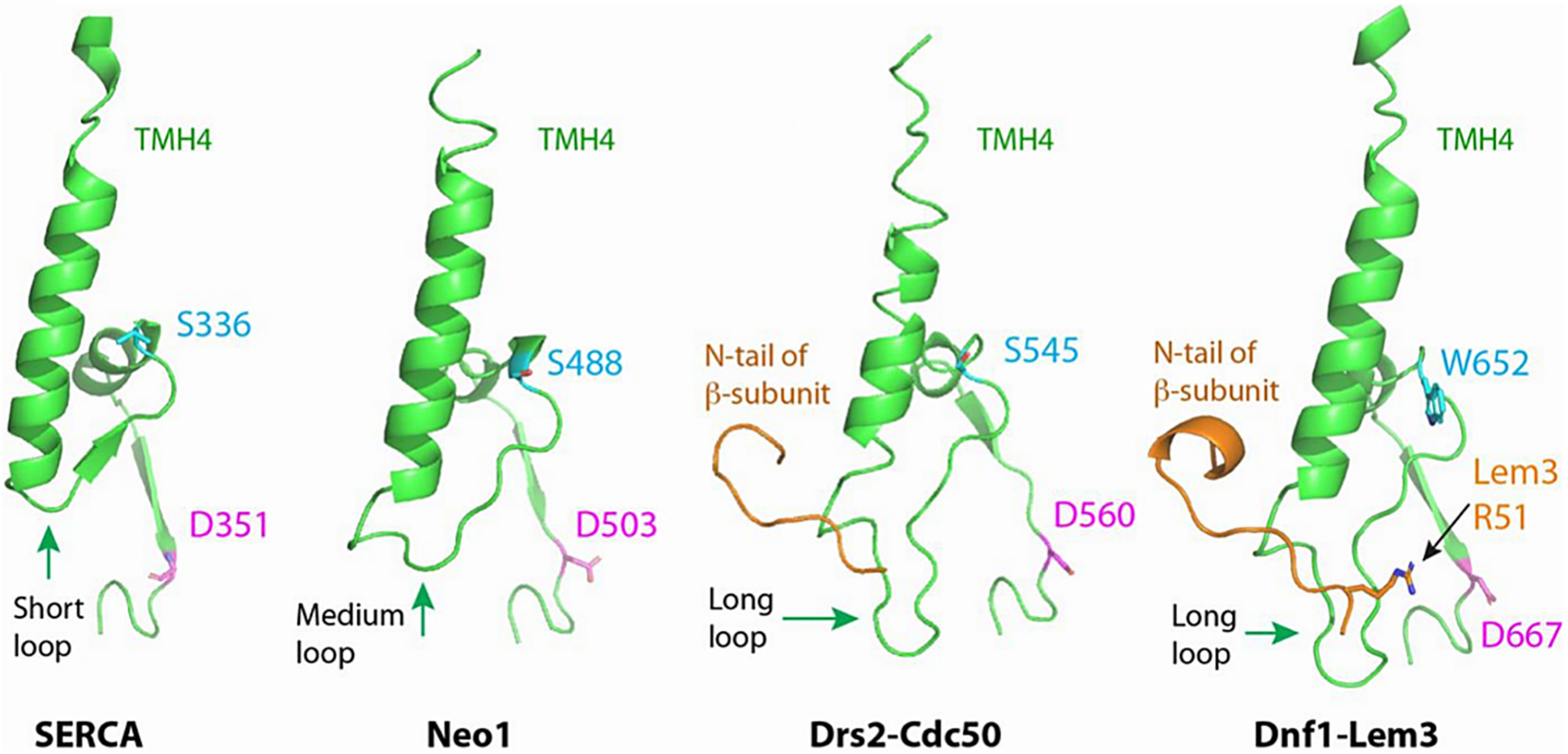


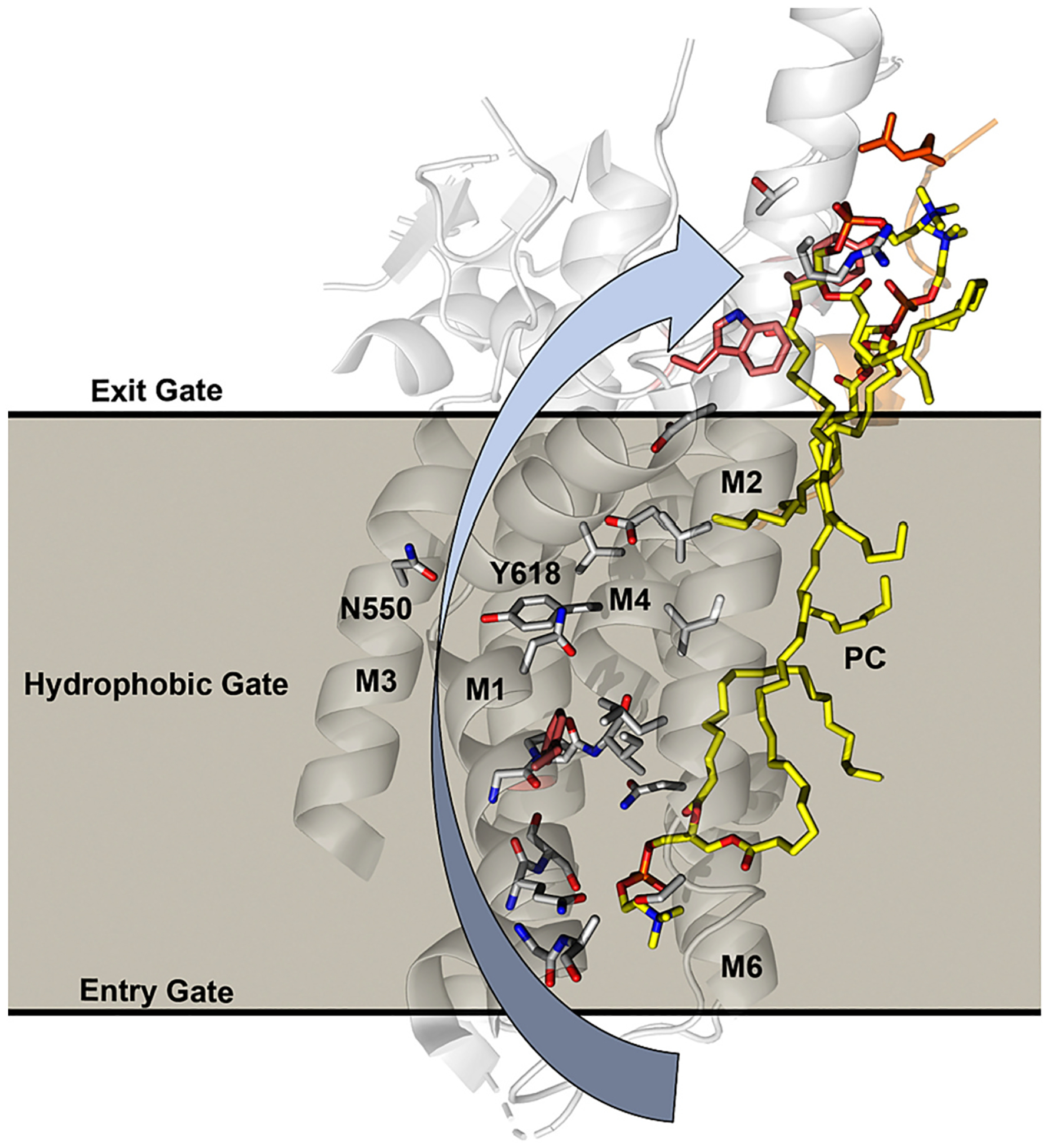

References
-
- Alberts B, et al. , Molecular Biology of the Cell, 7th ed., W.W. Norton & Co., Chapters 10–12, 2022.
-
- Harayama T, Riezman H, Understanding the diversity of membrane lipid composition, Nat. Rev. Mol. Cell Biol 19 (5) (2018) 281–296. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
