

Regulation of β-cell death by ADP-ribosylhydrolase ARH3 via lipid signaling in insulitis
- PMID: 38383396
- PMCID: PMC10880366
- DOI: 10.1186/s12964-023-01437-1
Regulation of β-cell death by ADP-ribosylhydrolase ARH3 via lipid signaling in insulitis
Abstract
Background: Lipids are regulators of insulitis and β-cell death in type 1 diabetes development, but the underlying mechanisms are poorly understood. Here, we investigated how the islet lipid composition and downstream signaling regulate β-cell death.
Methods: We performed lipidomics using three models of insulitis: human islets and EndoC-βH1 β cells treated with the pro-inflammatory cytokines interlukine-1β and interferon-γ, and islets from pre-diabetic non-obese mice. We also performed mass spectrometry and fluorescence imaging to determine the localization of lipids and enzyme in islets. RNAi, apoptotic assay, and qPCR were performed to determine the role of a specific factor in lipid-mediated cytokine signaling.
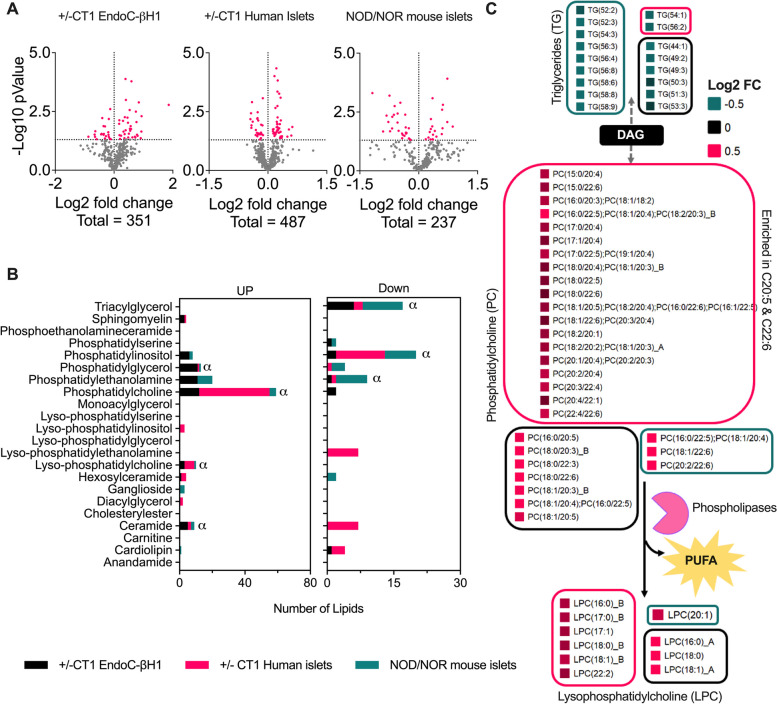
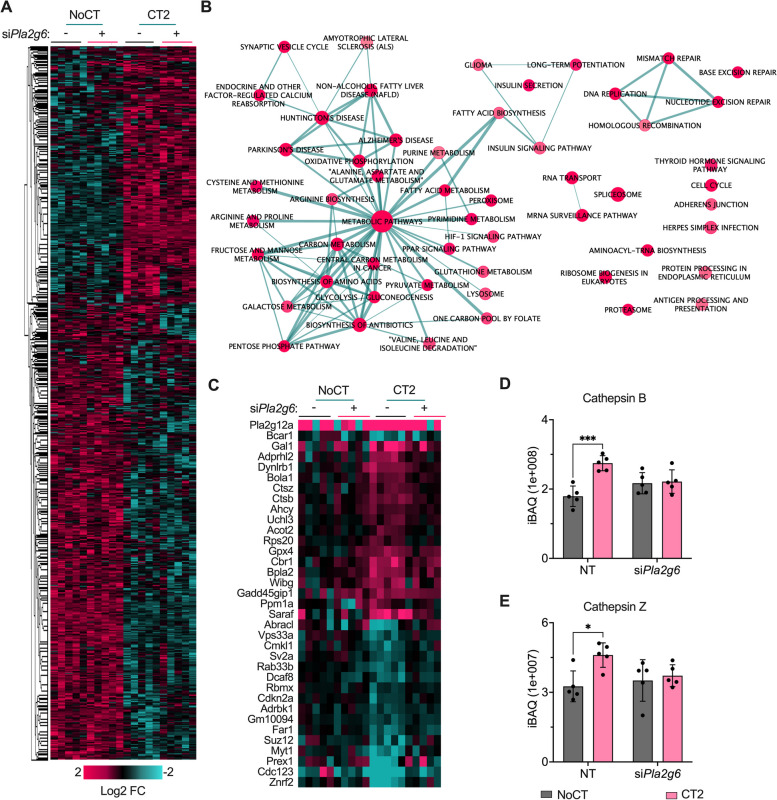
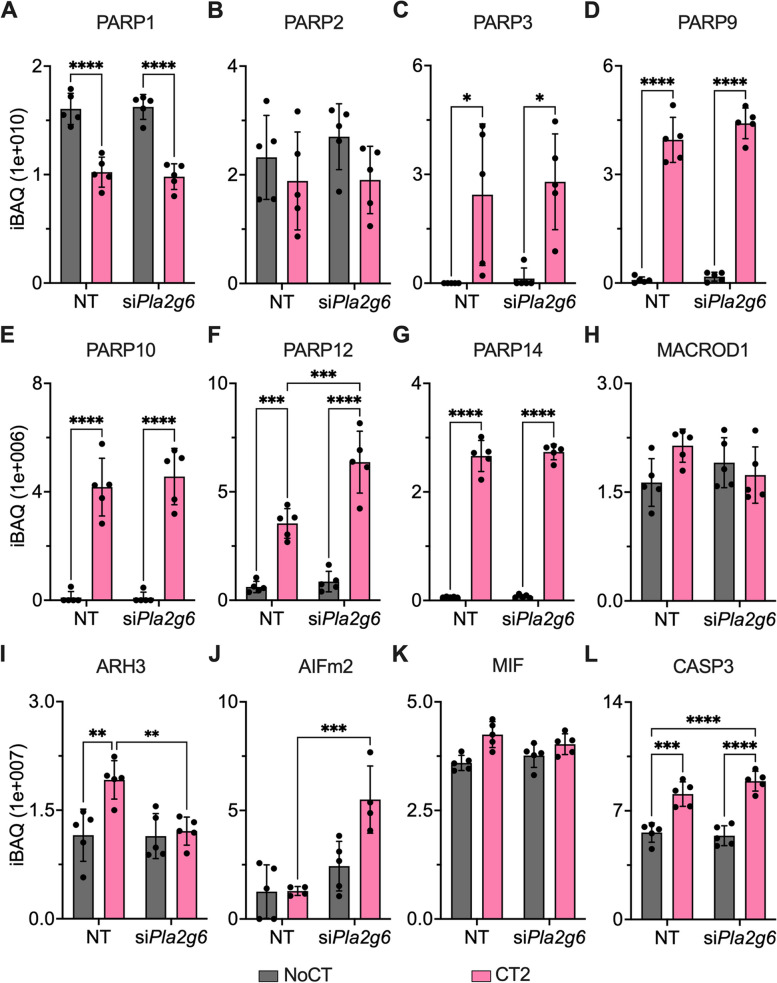
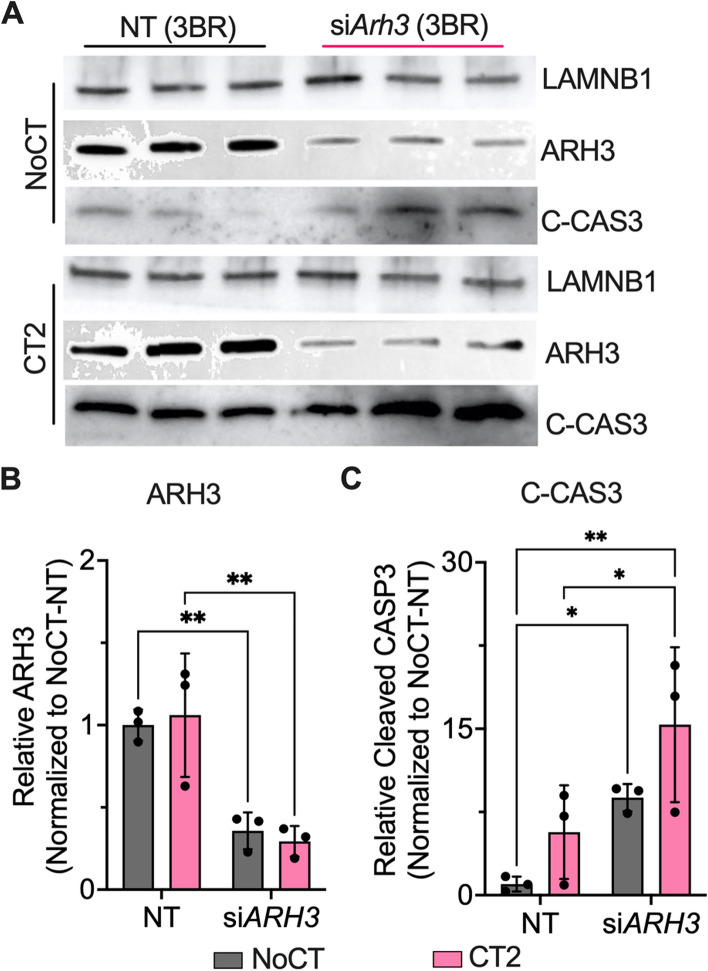
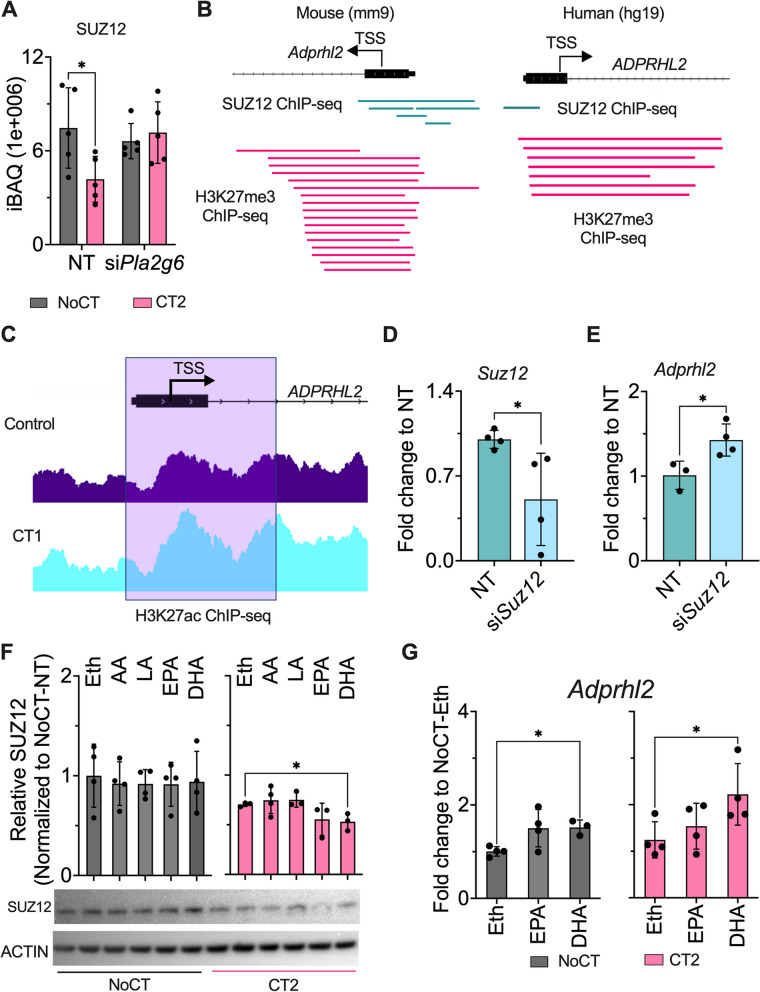
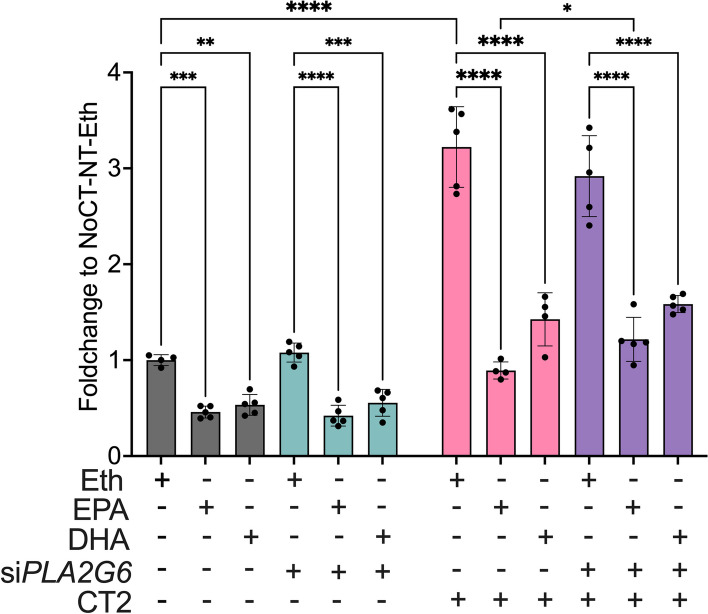
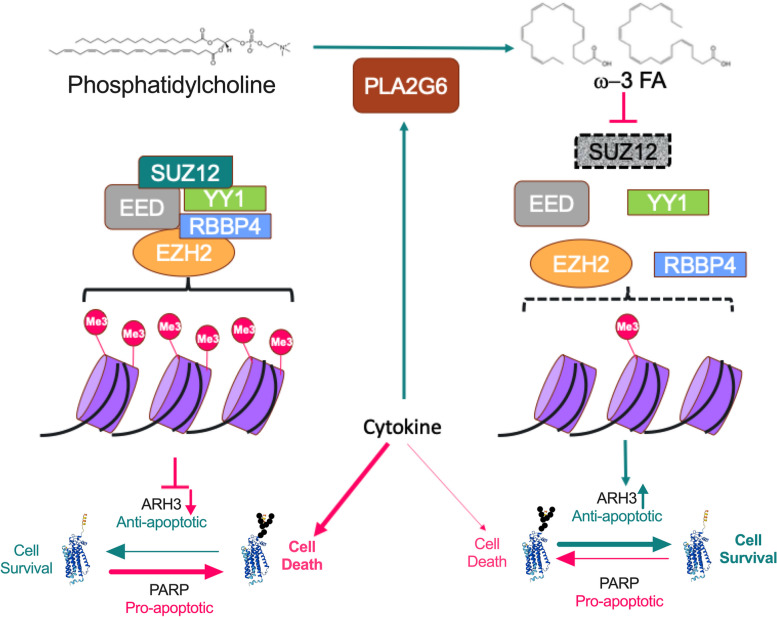
Results: Across all three models, lipidomic analyses showed a consistent increase of lysophosphatidylcholine species and phosphatidylcholines with polyunsaturated fatty acids and a reduction of triacylglycerol species. Imaging assays showed that phosphatidylcholines with polyunsaturated fatty acids and their hydrolyzing enzyme phospholipase PLA2G6 are enriched in islets. In downstream signaling, omega-3 fatty acids reduce cytokine-induced β-cell death by improving the expression of ADP-ribosylhydrolase ARH3. The mechanism involves omega-3 fatty acid-mediated reduction of the histone methylation polycomb complex PRC2 component Suz12, upregulating the expression of Arh3, which in turn decreases cell apoptosis.
Conclusions: Our data provide insights into the change of lipidomics landscape in β cells during insulitis and identify a protective mechanism by omega-3 fatty acids. Video Abstract.
© 2024. Battelle Memorial Institute.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
-
- Lamichhane S, Ahonen L, Dyrlund TS, Kemppainen E, Siljander H, Hyoty H, Ilonen J, Toppari J, Veijola R, Hyotylainen T, et al. Dynamics of plasma Lipidome in progression to islet autoimmunity and type 1 diabetes - type 1 diabetes prediction and prevention study (DIPP) Sci Rep. 2018;8:10635. doi: 10.1038/s41598-018-28907-8. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK060581/DK/NIDDK NIH HHS/United States
- R01 DK105588/DK/NIDDK NIH HHS/United States
- I01 BX001733/BX/BLRD VA/United States
- R01 DK127236/DK/NIDDK NIH HHS/United States
- P30 DK097512/DK/NIDDK NIH HHS/United States
- U01 DK127505/DK/NIDDK NIH HHS/United States
- R01 DK126444/DK/NIDDK NIH HHS/United States
- P30 DK020595/DK/NIDDK NIH HHS/United States
- UC4 DK104166/DK/NIDDK NIH HHS/United States
- P30 DK048520/DK/NIDDK NIH HHS/United States
- UC4 DK108101/DK/NIDDK NIH HHS/United States
- R01 DK093954/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
