

Structure of the recombinant RNA polymerase from African Swine Fever Virus
- PMID: 38383525
- PMCID: PMC10881513
- DOI: 10.1038/s41467-024-45842-7
Structure of the recombinant RNA polymerase from African Swine Fever Virus
Abstract
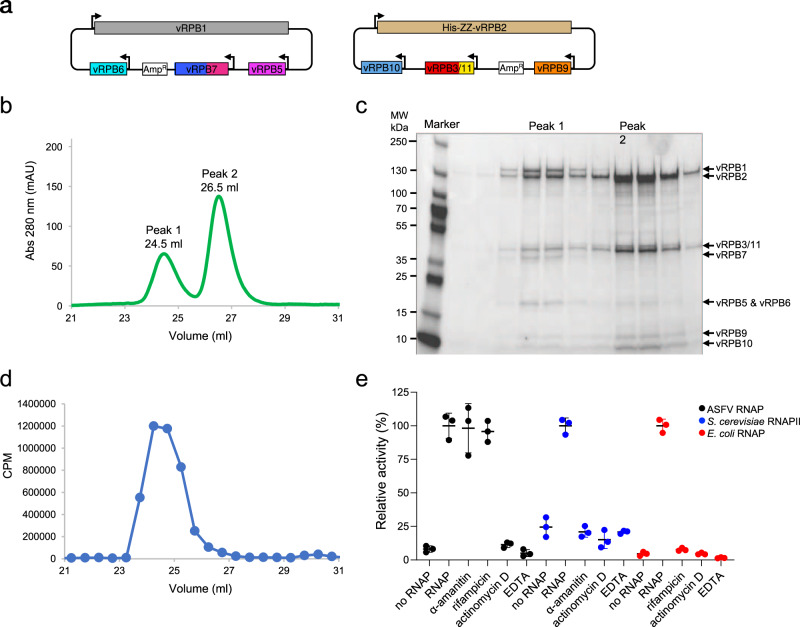
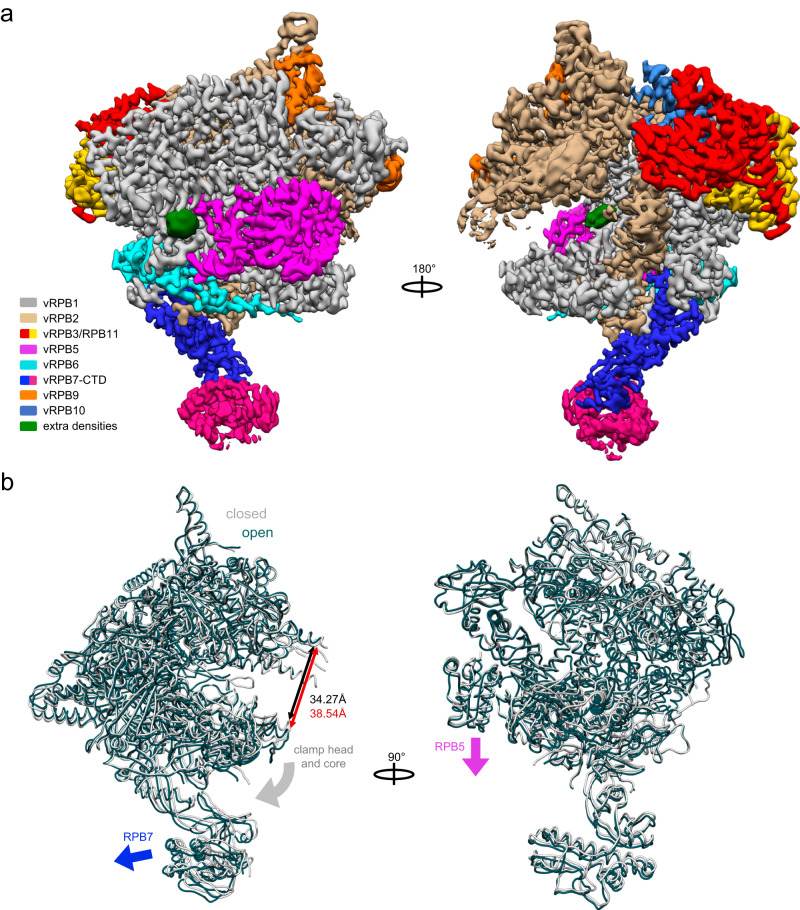
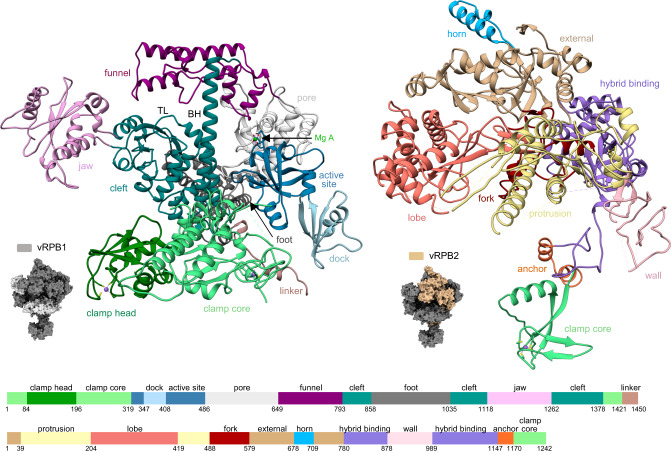
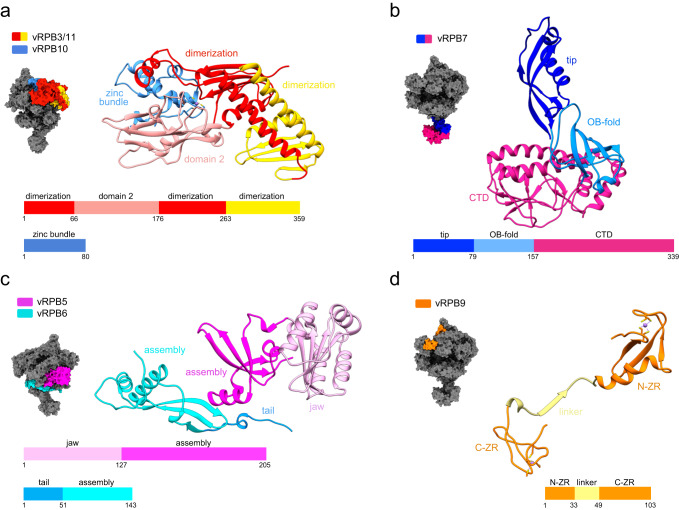
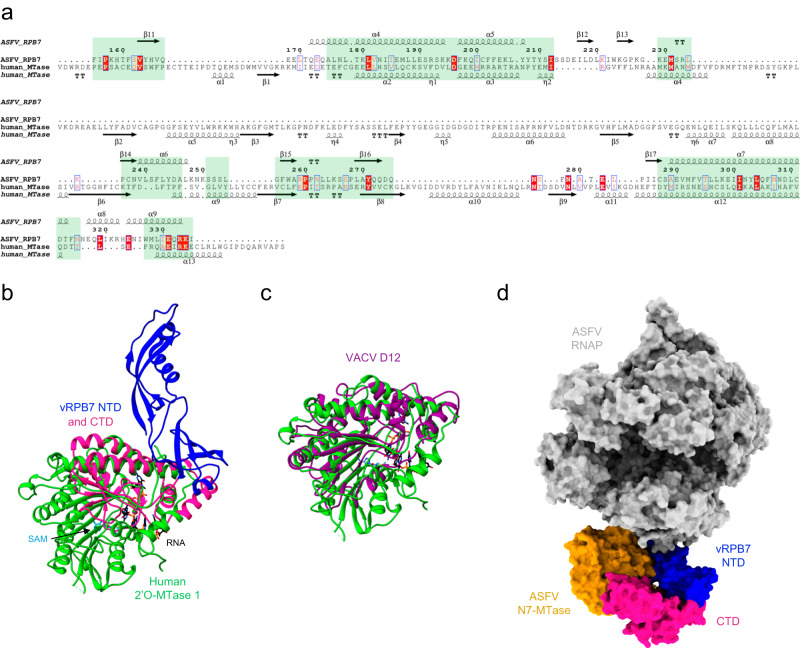
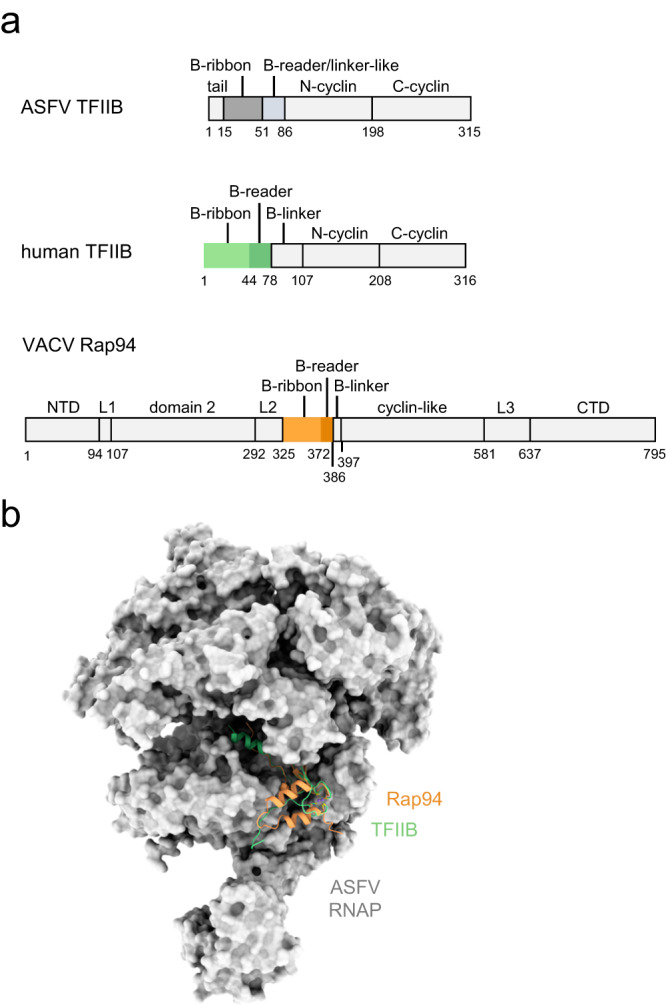
African Swine Fever Virus is a Nucleo-Cytoplasmic Large DNA Virus that causes an incurable haemorrhagic fever in pigs with a high impact on global food security. ASFV replicates in the cytoplasm of the infected cell and encodes its own transcription machinery that is independent of cellular factors, however, not much is known about how this system works at a molecular level. Here, we present methods to produce recombinant ASFV RNA polymerase, functional assays to screen for inhibitors, and high-resolution cryo-electron microscopy structures of the ASFV RNAP in different conformational states. The ASFV RNAP bears a striking resemblance to RNAPII with bona fide homologues of nine of its twelve subunits. Key differences include the fusion of the ASFV assembly platform subunits RPB3 and RPB11, and an unusual C-terminal domain of the stalk subunit vRPB7 that is related to the eukaryotic mRNA cap 2´-O-methyltransferase 1. Despite the high degree of structural conservation with cellular RNA polymerases, the ASFV RNAP is resistant to the inhibitors rifampicin and alpha-amanitin. The cryo-EM structures and fully recombinant RNAP system together provide an important tool for the design, development, and screening of antiviral drugs in a low biosafety containment environment.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Transcription termination and readthrough in African swine fever virus.Front Immunol. 2024 Mar 13;15:1350267. doi: 10.3389/fimmu.2024.1350267. eCollection 2024. Front Immunol. 2024. PMID: 38545109 Free PMC article.
-
The African Swine Fever Virus Transcriptome.J Virol. 2020 Apr 16;94(9):e00119-20. doi: 10.1128/JVI.00119-20. Print 2020 Apr 16. J Virol. 2020. PMID: 32075923 Free PMC article.
-
DNA-Binding Properties of African Swine Fever Virus pA104R, a Histone-Like Protein Involved in Viral Replication and Transcription.J Virol. 2017 May 26;91(12):e02498-16. doi: 10.1128/JVI.02498-16. Print 2017 Jun 15. J Virol. 2017. PMID: 28381576 Free PMC article.
-
Evolution of viral DNA-dependent RNA polymerases.Virus Genes. 1995;11(2-3):271-84. doi: 10.1007/BF01728665. Virus Genes. 1995. PMID: 8828152 Review.
-
Research progress on the proteins involved in African swine fever virus infection and replication.Front Immunol. 2022 Jul 22;13:947180. doi: 10.3389/fimmu.2022.947180. eCollection 2022. Front Immunol. 2022. PMID: 35935977 Free PMC article. Review.
Cited by
-
Genetic Variations of African Swine Fever Virus: Major Challenges and Prospects.Viruses. 2024 Jun 4;16(6):913. doi: 10.3390/v16060913. Viruses. 2024. PMID: 38932205 Free PMC article. Review.
-
Transcription regulation of African swine fever virus: dual role of M1249L.Nat Commun. 2024 Nov 20;15(1):10058. doi: 10.1038/s41467-024-54461-1. Nat Commun. 2024. PMID: 39567541 Free PMC article.
-
Transcription termination and readthrough in African swine fever virus.Front Immunol. 2024 Mar 13;15:1350267. doi: 10.3389/fimmu.2024.1350267. eCollection 2024. Front Immunol. 2024. PMID: 38545109 Free PMC article.
-
Structural basis of RNA polymerase complexes in African swine fever virus.Nat Commun. 2025 Jan 8;16(1):501. doi: 10.1038/s41467-024-55683-z. Nat Commun. 2025. PMID: 39779680 Free PMC article.
-
Advances in African swine fever virus molecular biology and host interactions contributing to new tools for control.J Virol. 2025 Jun 17;99(6):e0093224. doi: 10.1128/jvi.00932-24. Epub 2025 May 9. J Virol. 2025. PMID: 40340396 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
