

Characterization of the Zinc Uptake Repressor (Zur) from Acinetobacter baumannii
- PMID: 38385972
- PMCID: PMC11019503
- DOI: 10.1021/acs.biochem.3c00679
Characterization of the Zinc Uptake Repressor (Zur) from Acinetobacter baumannii
Abstract
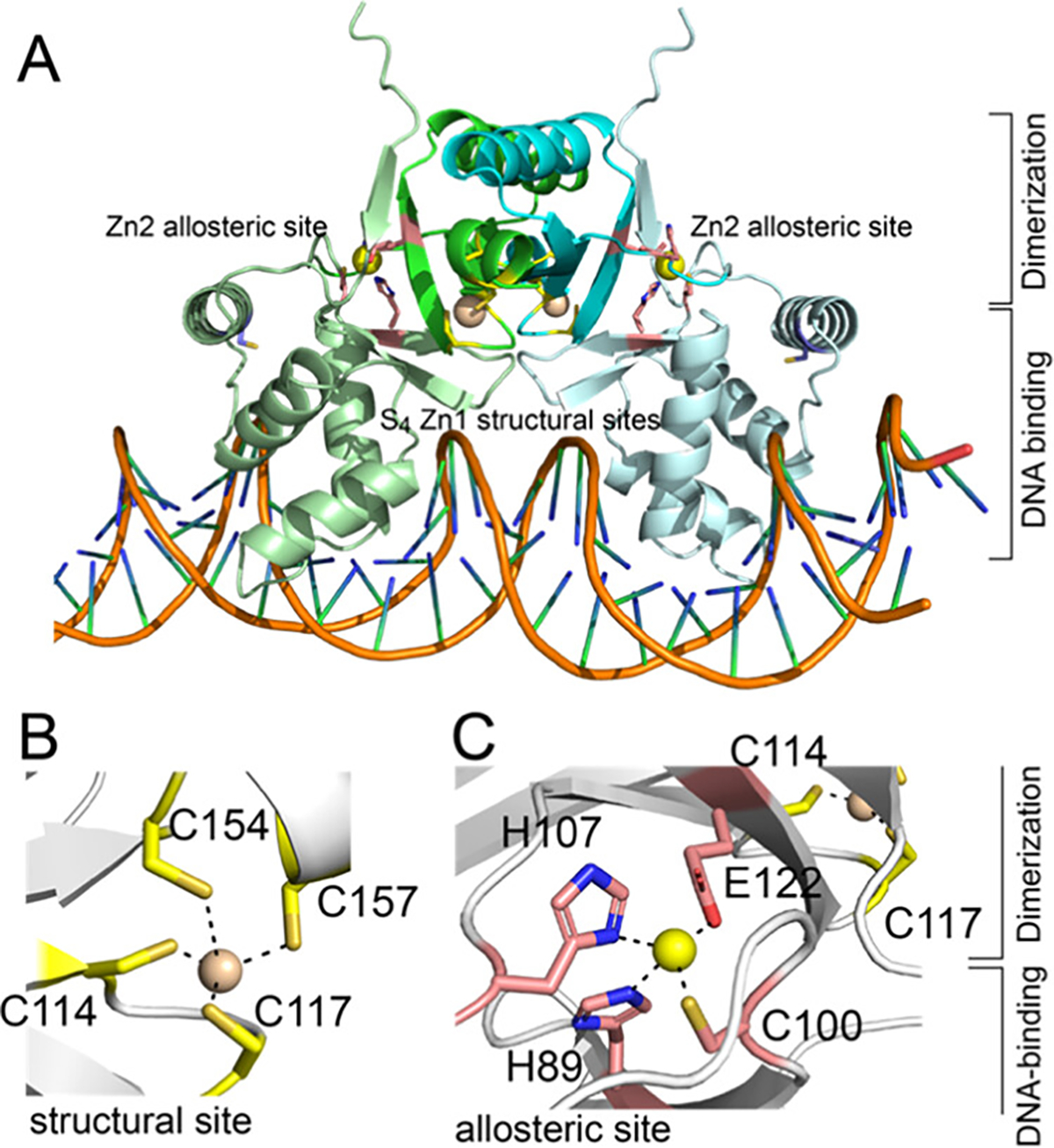
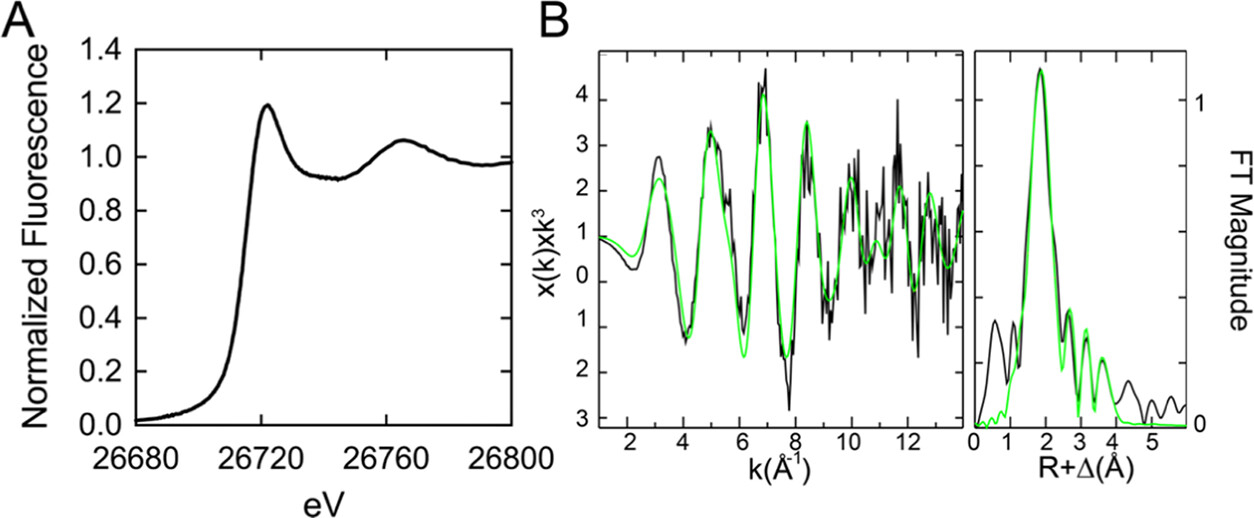
Bacterial cells tightly regulate the intracellular concentrations of essential transition metal ions by deploying a panel of metal-regulated transcriptional repressors and activators that bind to operator-promoter regions upstream of regulated genes. Like other zinc uptake regulator (Zur) proteins, Acinetobacter baumannii Zur represses transcription of its regulon when ZnII is replete and binds more weakly to DNA when ZnII is limiting. Previous studies established that Zur proteins are homodimeric and harbor at least two metal sites per protomer or four per dimer. CdII X-ray absorption spectroscopy (XAS) of the Cd2Zn2 AbZur metalloderivative with CdII bound to the allosteric sites reveals a S(N/O)3 first coordination shell. Site-directed mutagenesis suggests that H89 and C100 from the N-terminal DNA binding domain and H107 and E122 from the C-terminal dimerization domain comprise the regulatory metal site. KZn for this allosteric site is 6.0 (±2.2) × 1012 M-1 with a functional "division of labor" among the four metal ligands. N-terminal domain ligands H89 and C100 contribute far more to KZn than H107 and E122, while C100S AbZur uniquely fails to bind to DNA tightly as measured by an in vitro transcription assay. The heterotropic allosteric coupling free energy, ΔGc, is negative, consistent with a higher KZn for the AbZur-DNA complex and defining a bioavailable ZnII set-point of ≈6 × 10-14 M. Small-angle X-ray scattering (SAXS) experiments reveal that only the wild-type Zn homodimer undergoes allosteric switching, while the C100S AbZur fails to switch. These data collectively suggest that switching to a high affinity DNA-binding conformation involves a rotation/translation of one protomer relative to the other in a way that is dependent on the integrity of C100. We place these findings in the context of other Zur proteins and Fur family repressors more broadly.
Conflict of interest statement
The authors declare no competing financial interest.
Figures





Similar articles
-
Characterization of the metal receptor sites in Escherichia coli Zur, an ultrasensitive zinc(II) metalloregulatory protein.Biochemistry. 2001 Sep 4;40(35):10417-23. doi: 10.1021/bi0155448. Biochemistry. 2001. PMID: 11523983
-
Physical characterization of the manganese-sensing pneumococcal surface antigen repressor from Streptococcus pneumoniae.Biochemistry. 2013 Oct 29;52(43):7689-701. doi: 10.1021/bi401132w. Epub 2013 Oct 14. Biochemistry. 2013. PMID: 24067066 Free PMC article.
-
Elucidation of primary (alpha(3)N) and vestigial (alpha(5)) heavy metal-binding sites in Staphylococcus aureus pI258 CadC: evolutionary implications for metal ion selectivity of ArsR/SmtB metal sensor proteins.J Mol Biol. 2002 Jun 7;319(3):685-701. doi: 10.1016/S0022-2836(02)00299-1. J Mol Biol. 2002. PMID: 12054863
-
The SmtB/ArsR family of metalloregulatory transcriptional repressors: Structural insights into prokaryotic metal resistance.FEMS Microbiol Rev. 2003 Jun;27(2-3):131-43. doi: 10.1016/S0168-6445(03)00054-8. FEMS Microbiol Rev. 2003. PMID: 12829264 Review.
-
Bacterial zinc uptake regulator proteins and their regulons.Biochem Soc Trans. 2018 Aug 20;46(4):983-1001. doi: 10.1042/BST20170228. Epub 2018 Jul 31. Biochem Soc Trans. 2018. PMID: 30065104 Free PMC article. Review.
Cited by
-
Bacterial Metallostasis: Metal Sensing, Metalloproteome Remodeling, and Metal Trafficking.Chem Rev. 2024 Dec 25;124(24):13574-13659. doi: 10.1021/acs.chemrev.4c00264. Epub 2024 Dec 10. Chem Rev. 2024. PMID: 39658019 Free PMC article. Review.
-
The Coordination Chemistry of Two Peptidic Models of NFeoB and Core CFeoB Regions of FeoB Protein: Complexes of Fe(II), Mn(II), and Zn(II).Inorg Chem. 2025 Mar 17;64(10):5038-5052. doi: 10.1021/acs.inorgchem.4c05111. Epub 2025 Mar 6. Inorg Chem. 2025. PMID: 40048504 Free PMC article.
-
Metals in Motion: Understanding Labile Metal Pools in Bacteria.Biochemistry. 2025 Jan 21;64(2):329-345. doi: 10.1021/acs.biochem.4c00726. Epub 2025 Jan 5. Biochemistry. 2025. PMID: 39755956 Free PMC article. Review.
-
Coupling of zinc and GTP binding drives G-domain folding in Acinetobacter baumannii ZigA.Biophys J. 2024 Apr 16;123(8):979-991. doi: 10.1016/j.bpj.2024.03.010. Epub 2024 Mar 8. Biophys J. 2024. PMID: 38459695 Free PMC article.
References
-
- Waldron KJ; Rutherford JC; Ford D; Robinson NJ Metalloproteins and metal sensing. Nature 2009, 460, 823–830. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
