

Tumor-Associated Monocytes Reprogram CD8+ T Cells into Central Memory-Like Cells with Potent Antitumor Effects
- PMID: 38386350
- PMCID: PMC11040375
- DOI: 10.1002/advs.202304501
Tumor-Associated Monocytes Reprogram CD8+ T Cells into Central Memory-Like Cells with Potent Antitumor Effects
Abstract
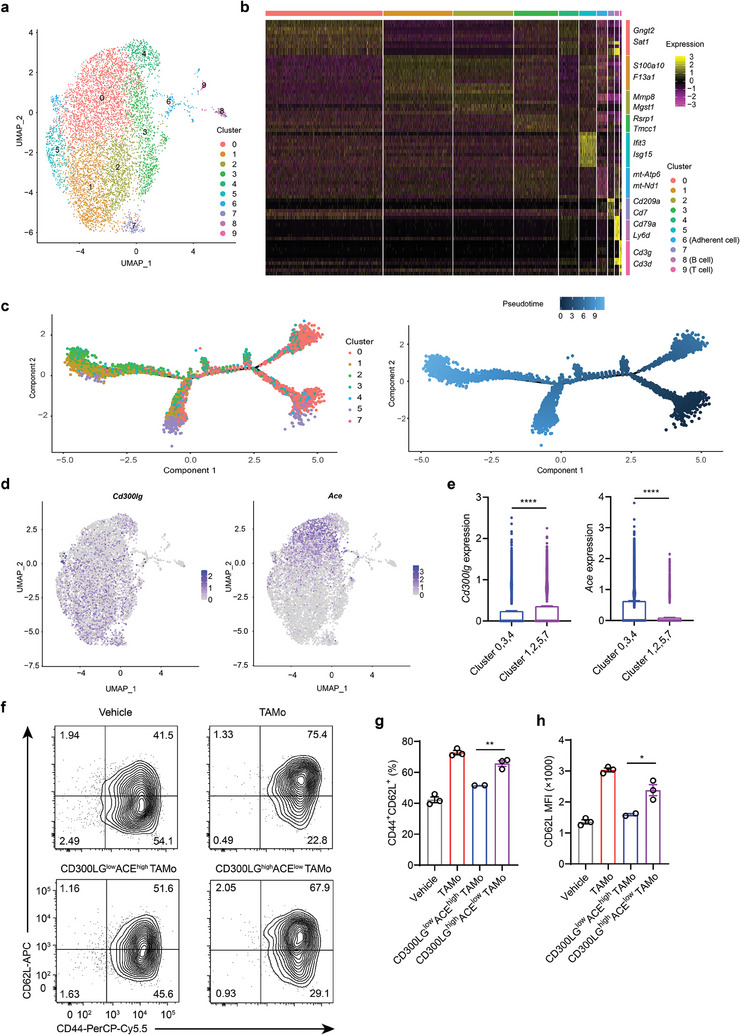
CD8+ T cells are critical for host antitumor responses, whereas persistent antigenic stimulation and excessive inflammatory signals lead to T cell dysfunction or exhaustion. Increasing early memory T cells can improve T cell persistence and empower T cell-mediated tumor eradication, especially for adoptive cancer immunotherapy. Here, it is reported that tumor-associated monocytes (TAMos) are highly correlated with the accumulation of CD8+ memory T cells in human cancers. Further analysis identifies that TAMos selectively reprogram CD8+ T cells into T central memory-like (TCM-like) cells with enhanced recall responses. L-NMMA, a pan nitric oxide synthase inhibitor, can mitigate TAMo-mediated inhibition of T cell proliferation without affecting TCM-like cell generation. Moreover, the modified T cells by TAMo exposure and L-NMMA treatment exhibit long-term persistence and elicit superior antitumor effects in vivo. Mechanistically, the transmembrane protein CD300LG is involved in TAMo-mediated TCM-like cell polarization in a cell-cell contact-dependent manner. Thus, the terminally differentiated TAMo subset (CD300LGhighACElow) mainly contributes to TCM-like cell development. Taken together, these findings establish the significance of TAMos in boosting T-cell antitumor immunity.
Keywords: CD300LG; T cell exhaustion; T central memory‐like cells; nitric oxide synthase; tumor‐associated monocytes (TAMos).
© 2024 The Authors. Advanced Science published by Wiley‐VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







Similar articles
-
Mimicking homeostatic proliferation in vitro generates T cells with high anti-tumor function in non-lymphopenic hosts.Cancer Immunol Immunother. 2013 Mar;62(3):503-15. doi: 10.1007/s00262-012-1350-1. Epub 2012 Sep 23. Cancer Immunol Immunother. 2013. PMID: 23001162 Free PMC article.
-
Central memory self/tumor-reactive CD8+ T cells confer superior antitumor immunity compared with effector memory T cells.Proc Natl Acad Sci U S A. 2005 Jul 5;102(27):9571-6. doi: 10.1073/pnas.0503726102. Epub 2005 Jun 24. Proc Natl Acad Sci U S A. 2005. PMID: 15980149 Free PMC article.
-
Transient stimulation expands superior antitumor T cells for adoptive therapy.JCI Insight. 2017 Jan 26;2(2):e89580. doi: 10.1172/jci.insight.89580. JCI Insight. 2017. PMID: 28138559 Free PMC article.
-
Memory T cells in cancer immunotherapy: which CD8 T-cell population provides the best protection against tumours?Tissue Antigens. 2008 Sep;72(3):187-94. doi: 10.1111/j.1399-0039.2008.01088.x. Epub 2008 Jul 9. Tissue Antigens. 2008. PMID: 18627571 Review.
-
T-memory cells against cancer: Remembering the enemy.Cell Immunol. 2019 Apr;338:27-31. doi: 10.1016/j.cellimm.2019.03.002. Epub 2019 Mar 16. Cell Immunol. 2019. PMID: 30928016 Review.
Cited by
-
Machine learning and single-cell analysis uncover distinctive characteristics of CD300LG within the TNBC immune microenvironment: experimental validation.Clin Exp Med. 2025 May 17;25(1):167. doi: 10.1007/s10238-025-01690-3. Clin Exp Med. 2025. PMID: 40382513 Free PMC article.
-
Longitudinal Monitoring of T cell Dynamics in Metastatic Breast Cancer via a Remote Diagnostic Implant.Immunomedicine. 2024 Dec;4(2):10.1002/imed.70000. doi: 10.1002/imed.70000. Epub 2025 Jun 9. Immunomedicine. 2024. PMID: 40880982
-
The efficacy of plasma exosomal miRNAs as predictive biomarkers for PD-1 blockade plus chemotherapy in gastric cancer.Transl Cancer Res. 2024 Nov 30;13(11):6336-6346. doi: 10.21037/tcr-24-2151. Epub 2024 Nov 27. Transl Cancer Res. 2024. PMID: 39697742 Free PMC article.
-
Single-cell mitophagy patterns within the tumor microenvironment modulate intercellular communication, impacting the progression and prognosis of hepatocellular carcinoma.Front Immunol. 2025 Jan 6;15:1448878. doi: 10.3389/fimmu.2024.1448878. eCollection 2024. Front Immunol. 2025. PMID: 39835122 Free PMC article.
-
Microwave Ablation Combined with Flt3L Provokes Tumor-Specific Memory CD8+ T Cells-Mediated Antitumor Immunity in Response to PD-1 Blockade.Adv Sci (Weinh). 2025 Jan;12(4):e2413181. doi: 10.1002/advs.202413181. Epub 2024 Dec 4. Adv Sci (Weinh). 2025. PMID: 39629989 Free PMC article.
References
-
- Fraietta J. A., Nobles C. L., Sammons M. A., Lundh S., Carty S. A., Reich T. J., Cogdill A. P., Morrissette J. J. D., DeNizio J. E., Reddy S., Hwang Y., Gohil M., Kulikovskaya I., Nazimuddin F., Gupta M., Chen F., Everett J. K., Alexander K. A., Lin‐Shiao E., Gee M. H., Liu X., Young R. M., Ambrose D., Wang Y., Xu J., Jordan M. S., Marcucci K. T., Levine B. L., Garcia K. C., Zhao Y., Nature 2018, 558, 307. - PMC - PubMed
-
- Mo F., Yu Z., Li P., Oh J., Spolski R., Zhao L., Glassman C. R., Yamamoto T. N., Chen Y., Golebiowski F. M., Hermans D., Majri‐Morrison S., Picton L. K., Liao W., Ren M., Zhuang X., Mitra S., Lin J. X., Gattinoni L., Powell J. D., Restifo N. P., Garcia K. C., Leonard W. J., Nature 2021, 597, 544. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous