

Denaturing purifications demonstrate that PRC2 and other widely reported chromatin proteins do not appear to bind directly to RNA in vivo
- PMID: 38387462
- PMCID: PMC10997485
- DOI: 10.1016/j.molcel.2024.01.026
Denaturing purifications demonstrate that PRC2 and other widely reported chromatin proteins do not appear to bind directly to RNA in vivo
Abstract
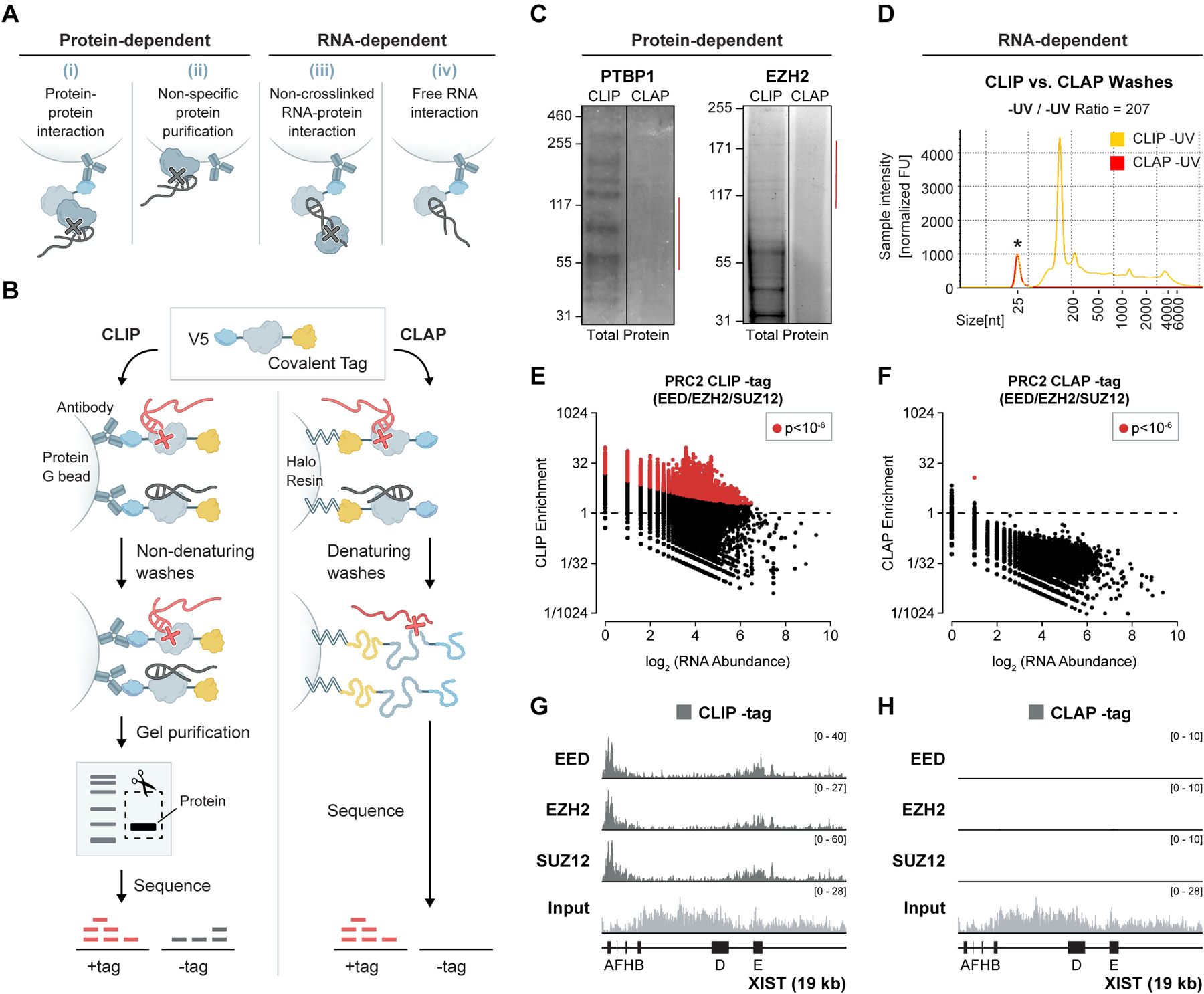
Polycomb repressive complex 2 (PRC2) is reported to bind to many RNAs and has become a central player in reports of how long non-coding RNAs (lncRNAs) regulate gene expression. Yet, there is a growing discrepancy between the biochemical evidence supporting specific lncRNA-PRC2 interactions and functional evidence demonstrating that PRC2 is often dispensable for lncRNA function. Here, we revisit the evidence supporting RNA binding by PRC2 and show that many reported interactions may not occur in vivo. Using denaturing purification of in vivo crosslinked RNA-protein complexes in human and mouse cell lines, we observe a loss of detectable RNA binding to PRC2 and chromatin-associated proteins previously reported to bind RNA (CTCF, YY1, and others), despite accurately mapping bona fide RNA-binding sites across others (SPEN, TET2, and others). Taken together, these results argue for a critical re-evaluation of the broad role of RNA binding to orchestrate various chromatin regulatory mechanisms.
Keywords: CLIP; PRC2; RNA; RNA-binding proteins; chromatin.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests M.R.B., W.G.W., and M.G. are inventors on a patent covering the CLAP method.
Figures





Similar articles
-
Interactions between JARID2 and noncoding RNAs regulate PRC2 recruitment to chromatin.Mol Cell. 2014 Jan 23;53(2):290-300. doi: 10.1016/j.molcel.2013.11.012. Epub 2013 Dec 26. Mol Cell. 2014. PMID: 24374312 Free PMC article.
-
Association of the Long Non-coding RNA Steroid Receptor RNA Activator (SRA) with TrxG and PRC2 Complexes.PLoS Genet. 2015 Oct 23;11(10):e1005615. doi: 10.1371/journal.pgen.1005615. eCollection 2015 Oct. PLoS Genet. 2015. PMID: 26496121 Free PMC article.
-
Cryptic RNA-binding by PRC2 components EZH2 and SUZ12.RNA Biol. 2015;12(9):959-65. doi: 10.1080/15476286.2015.1069463. RNA Biol. 2015. PMID: 26177152 Free PMC article.
-
The control of polycomb repressive complexes by long noncoding RNAs.Wiley Interdiscip Rev RNA. 2021 Nov;12(6):e1657. doi: 10.1002/wrna.1657. Epub 2021 Apr 16. Wiley Interdiscip Rev RNA. 2021. PMID: 33861025 Free PMC article. Review.
-
The recruitment of chromatin modifiers by long noncoding RNAs: lessons from PRC2.RNA. 2015 Dec;21(12):2007-22. doi: 10.1261/rna.053918.115. RNA. 2015. PMID: 26574518 Free PMC article. Review.
Cited by
-
What's in a name: the multifaceted function of DNA- and RNA-binding proteins in T cell responses.FEBS J. 2025 Apr;292(8):1853-1867. doi: 10.1111/febs.17273. Epub 2024 Sep 20. FEBS J. 2025. PMID: 39304985 Free PMC article. Review.
-
A Compendium of G-Flipon Biological Functions That Have Experimental Validation.Int J Mol Sci. 2024 Sep 25;25(19):10299. doi: 10.3390/ijms251910299. Int J Mol Sci. 2024. PMID: 39408629 Free PMC article. Review.
-
Histone deacetylase-1 is required for epigenome stability in Neurospora crassa.bioRxiv [Preprint]. 2025 Jan 21:2025.01.17.633486. doi: 10.1101/2025.01.17.633486. bioRxiv. 2025. PMID: 39896537 Free PMC article. Preprint.
-
Intertwining roles of R-loops and G-quadruplexes in DNA repair, transcription and genome organization.Nat Cell Biol. 2024 Jul;26(7):1025-1036. doi: 10.1038/s41556-024-01437-4. Epub 2024 Jun 24. Nat Cell Biol. 2024. PMID: 38914786 Free PMC article. Review.
-
An RNA-centric view of transcription and genome organization.Mol Cell. 2024 Oct 3;84(19):3627-3643. doi: 10.1016/j.molcel.2024.08.021. Mol Cell. 2024. PMID: 39366351 Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
