

Euglena's atypical respiratory chain adapts to the discoidal cristae and flexible metabolism
- PMID: 38388527
- PMCID: PMC10884005
- DOI: 10.1038/s41467-024-46018-z
Euglena's atypical respiratory chain adapts to the discoidal cristae and flexible metabolism
Abstract
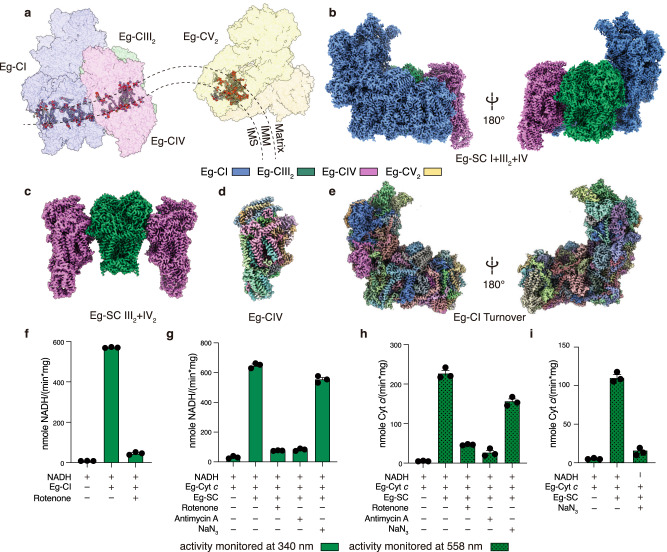
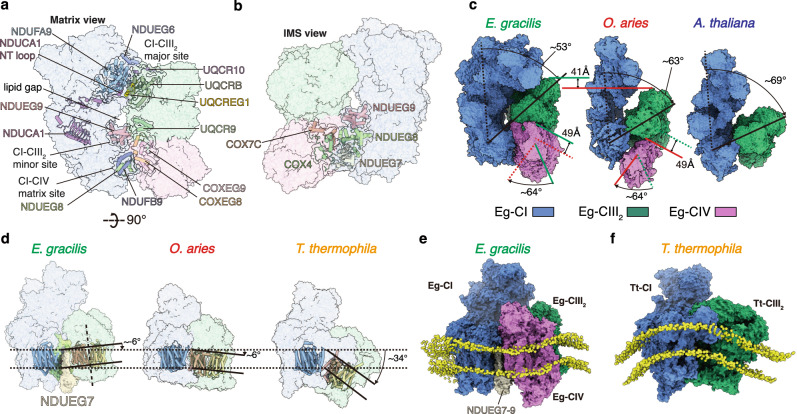
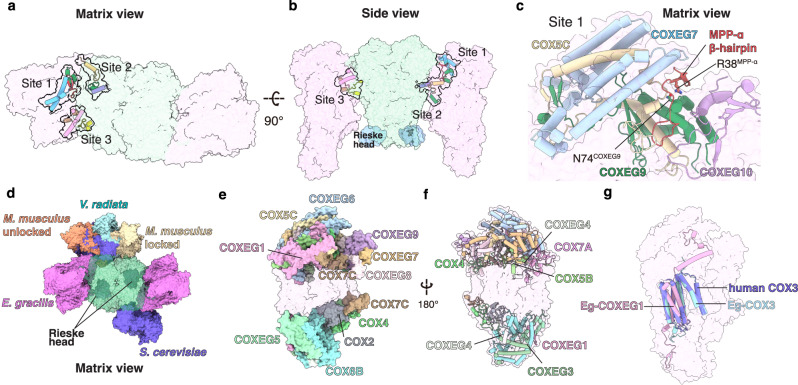
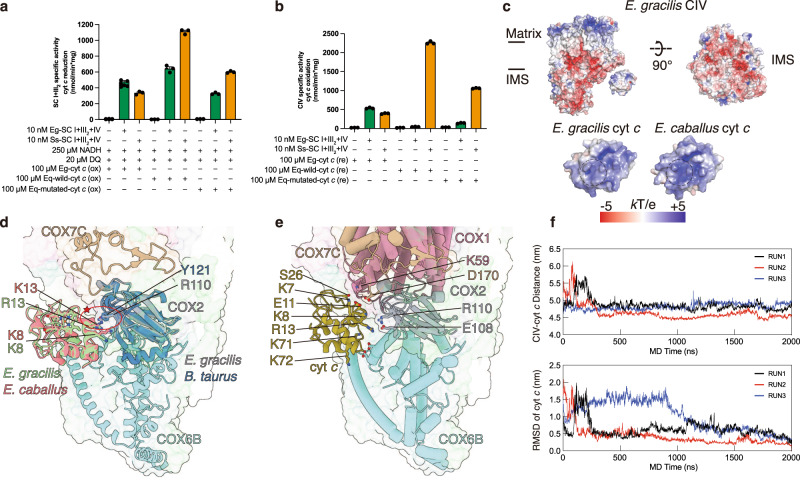
Euglena gracilis, a model organism of the eukaryotic supergroup Discoba harbouring also clinically important parasitic species, possesses diverse metabolic strategies and an atypical electron transport chain. While structures of the electron transport chain complexes and supercomplexes of most other eukaryotic clades have been reported, no similar structure is currently available for Discoba, limiting the understandings of its core metabolism and leaving a gap in the evolutionary tree of eukaryotic bioenergetics. Here, we report high-resolution cryo-EM structures of Euglena's respirasome I + III2 + IV and supercomplex III2 + IV2. A previously unreported fatty acid synthesis domain locates on the tip of complex I's peripheral arm, providing a clear picture of its atypical subunit composition identified previously. Individual complexes are re-arranged in the respirasome to adapt to the non-uniform membrane curvature of the discoidal cristae. Furthermore, Euglena's conformationally rigid complex I is deactivated by restricting ubiquinone's access to its substrate tunnel. Our findings provide structural insights for therapeutic developments against euglenozoan parasite infections.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Structures of Tetrahymena thermophila respiratory megacomplexes on the tubular mitochondrial cristae.Nat Commun. 2023 May 29;14(1):2542. doi: 10.1038/s41467-023-38158-5. Nat Commun. 2023. PMID: 37248254 Free PMC article.
-
Supramolecular associations between atypical oxidative phosphorylation complexes of Euglena gracilis.J Bioenerg Biomembr. 2021 Jun;53(3):351-363. doi: 10.1007/s10863-021-09882-8. Epub 2021 Mar 1. J Bioenerg Biomembr. 2021. PMID: 33646522 Free PMC article.
-
The atypical subunit composition of respiratory complexes I and IV is associated with original extra structural domains in Euglena gracilis.Sci Rep. 2018 Jun 26;8(1):9698. doi: 10.1038/s41598-018-28039-z. Sci Rep. 2018. PMID: 29946152 Free PMC article.
-
The road to the structure of the mitochondrial respiratory chain supercomplex.Biochem Soc Trans. 2020 Apr 29;48(2):621-629. doi: 10.1042/BST20190930. Biochem Soc Trans. 2020. PMID: 32311046 Free PMC article. Review.
-
Clarifying the supercomplex: the higher-order organization of the mitochondrial electron transport chain.Nat Struct Mol Biol. 2017 Oct 5;24(10):800-808. doi: 10.1038/nsmb.3460. Nat Struct Mol Biol. 2017. PMID: 28981073 Review.
Cited by
-
Biochemical and Functional Profiling of Thioredoxin-Dependent Cytosolic GPX-like Proteins in Euglena gracilis.Biomolecules. 2024 Jun 27;14(7):765. doi: 10.3390/biom14070765. Biomolecules. 2024. PMID: 39062479 Free PMC article.
-
Plant supercomplex I + III2 structure and function: implications for the growing field.Biochem Soc Trans. 2024 Aug 28;52(4):1647-1659. doi: 10.1042/BST20230947. Biochem Soc Trans. 2024. PMID: 39177070 Free PMC article. Review.
-
Integrated proteome and pangenome analysis revealed the variation of microalga Isochrysis galbana and associated bacterial community to 2,6-Di-tert-butyl-p-cresol (BHT) stress.World J Microbiol Biotechnol. 2024 Oct 24;40(11):364. doi: 10.1007/s11274-024-04171-z. World J Microbiol Biotechnol. 2024. PMID: 39446252
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
