

Structured 3' UTRs destabilize mRNAs in plants
- PMID: 38388963
- PMCID: PMC10885604
- DOI: 10.1186/s13059-024-03186-x
Structured 3' UTRs destabilize mRNAs in plants
Abstract
Background: RNA secondary structure (RSS) can influence the regulation of transcription, RNA processing, and protein synthesis, among other processes. 3' untranslated regions (3' UTRs) of mRNA also hold the key for many aspects of gene regulation. However, there are often contradictory results regarding the roles of RSS in 3' UTRs in gene expression in different organisms and/or contexts.
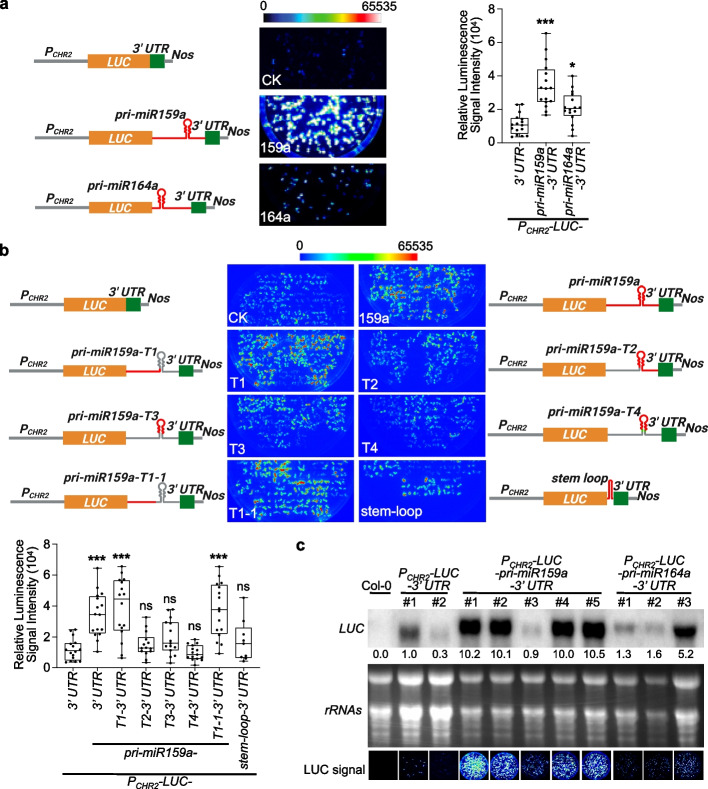
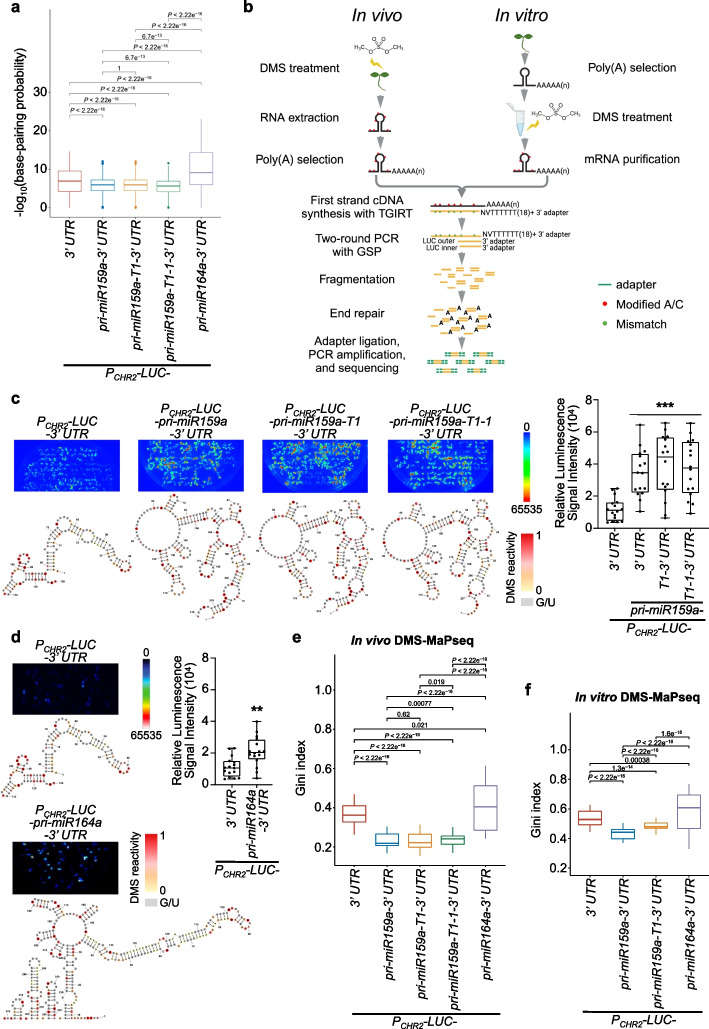
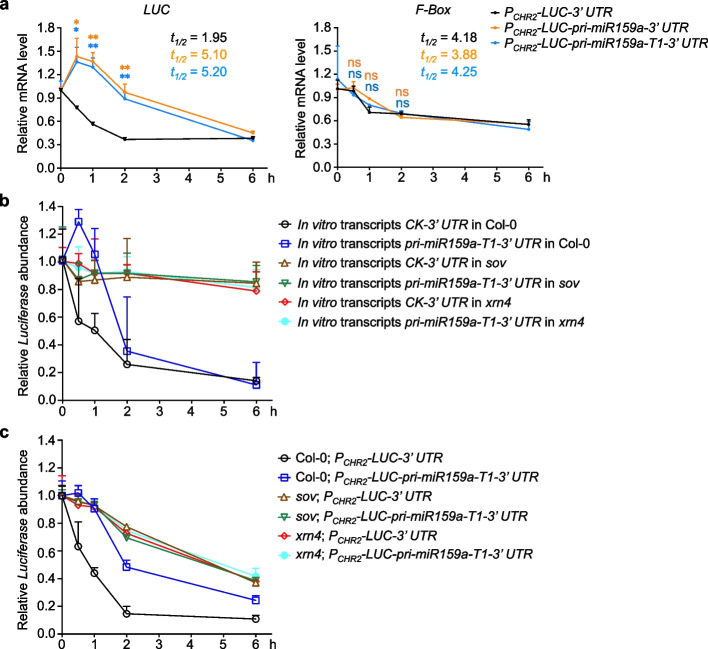
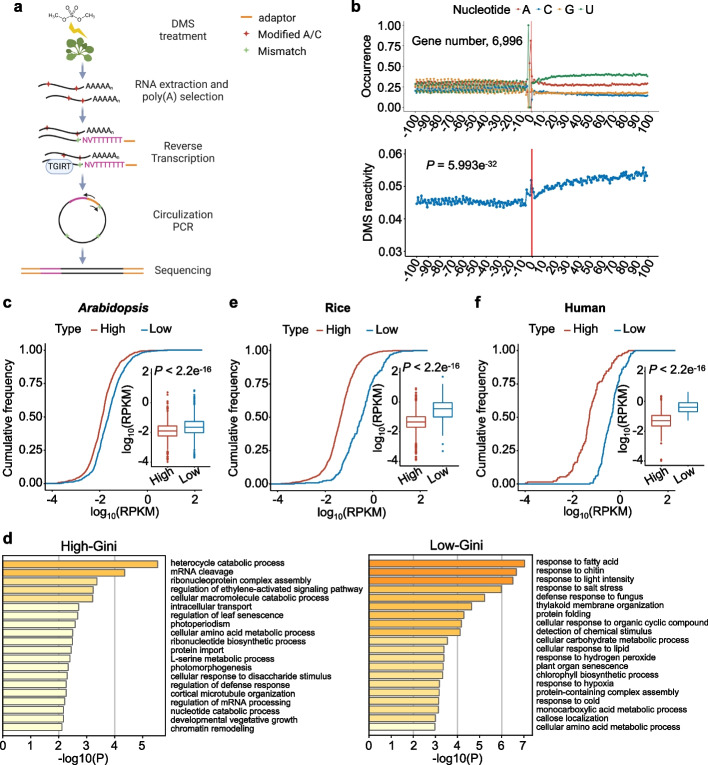
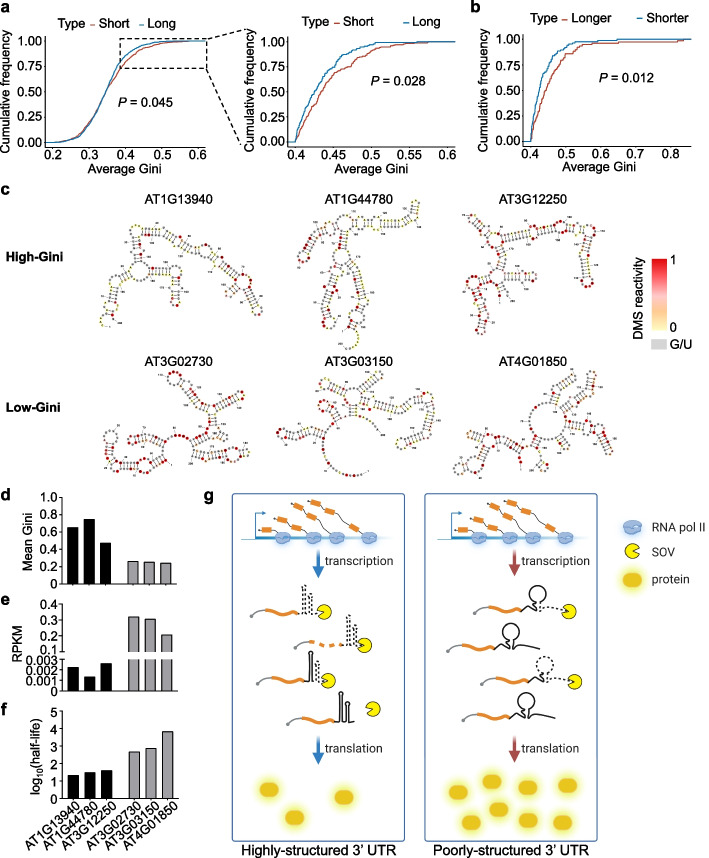
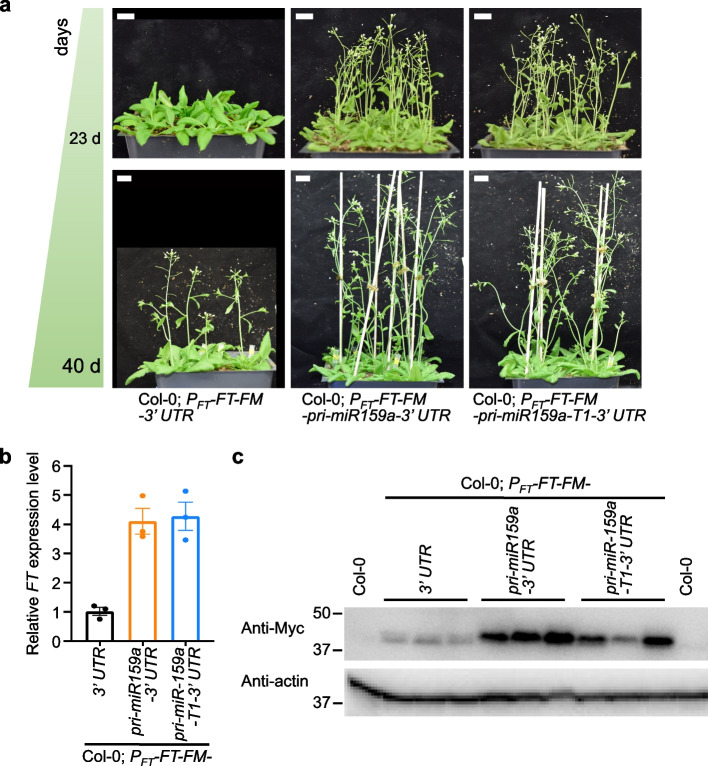
Results: Here, we incidentally observe that the primary substrate of miR159a (pri-miR159a), when embedded in a 3' UTR, could promote mRNA accumulation. The enhanced expression is attributed to the earlier polyadenylation of the transcript within the hybrid pri-miR159a-3' UTR and, resultantly, a poorly structured 3' UTR. RNA decay assays indicate that poorly structured 3' UTRs could promote mRNA stability, whereas highly structured 3' UTRs destabilize mRNA in vivo. Genome-wide DMS-MaPseq also reveals the prevailing inverse relationship between 3' UTRs' RSS and transcript accumulation in the transcriptomes of Arabidopsis, rice, and even human. Mechanistically, transcripts with highly structured 3' UTRs are preferentially degraded by 3'-5' exoribonuclease SOV and 5'-3' exoribonuclease XRN4, leading to decreased expression in Arabidopsis. Finally, we engineer different structured 3' UTRs to an endogenous FT gene and alter the FT-regulated flowering time in Arabidopsis.
Conclusions: We conclude that highly structured 3' UTRs typically cause reduced accumulation of the harbored transcripts in Arabidopsis. This pattern extends to rice and even mammals. Furthermore, our study provides a new strategy of engineering the 3' UTRs' RSS to modify plant traits in agricultural production and mRNA stability in biotechnology.
Keywords: 3′ UTR; 3′ end target-specific DMS-MaPseq; DIM-2P-seq; RNA secondary structure (RSS); mRNA stability.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
The role of secondary structures in the functioning of 3' untranslated regions of mRNA: A review of functions of 3' UTRs' secondary structures and hypothetical involvement of secondary structures in cytoplasmic polyadenylation in Drosophila.Bioessays. 2024 Mar;46(3):e2300099. doi: 10.1002/bies.202300099. Epub 2023 Dec 31. Bioessays. 2024. PMID: 38161240 Free PMC article. Review.
-
Dehydration stress extends mRNA 3' untranslated regions with noncoding RNA functions in Arabidopsis.Genome Res. 2017 Aug;27(8):1427-1436. doi: 10.1101/gr.218669.116. Epub 2017 May 18. Genome Res. 2017. PMID: 28522613 Free PMC article.
-
Evidence that XRN4, an Arabidopsis homolog of exoribonuclease XRN1, preferentially impacts transcripts with certain sequences or in particular functional categories.RNA. 2011 Mar;17(3):501-11. doi: 10.1261/rna.2467911. Epub 2011 Jan 11. RNA. 2011. PMID: 21224377 Free PMC article.
-
Regulation of co-translational mRNA decay by PAP and DXO1 in Arabidopsis.BMC Plant Biol. 2025 Feb 18;25(1):223. doi: 10.1186/s12870-025-06195-5. BMC Plant Biol. 2025. PMID: 39966730 Free PMC article.
-
Regulation by 3'-Untranslated Regions.Annu Rev Genet. 2017 Nov 27;51:171-194. doi: 10.1146/annurev-genet-120116-024704. Epub 2017 Aug 30. Annu Rev Genet. 2017. PMID: 28853924 Review.
Cited by
-
Unveiling RNA structure-mediated regulations of RNA stability in wheat.Nat Commun. 2024 Nov 20;15(1):10042. doi: 10.1038/s41467-024-54172-7. Nat Commun. 2024. PMID: 39567481 Free PMC article.
-
Quantifying the Evolution of SNPs That Affect RNA Secondary Structure in Arabidopsis thaliana Genes.Mol Biol Evol. 2025 Jun 4;42(6):msaf126. doi: 10.1093/molbev/msaf126. Mol Biol Evol. 2025. PMID: 40492874 Free PMC article.
-
Identification of RppSLN from an Elite Landrace: A Major Locus Conferring Resistance to Southern Corn Rust in Maize (Zea mays L.).Plants (Basel). 2024 Nov 16;13(22):3227. doi: 10.3390/plants13223227. Plants (Basel). 2024. PMID: 39599436 Free PMC article.
References
-
- Gosai SJ, Foley SW, Wang D, Silverman IM, Selamoglu N, Nelson AD, Beilstein MA, Daldal F, Deal RB, Gregory BD. Global analysis of the RNA-protein interaction and RNA secondary structure landscapes of the Arabidopsis nucleus. Mol Cell. 2015;57:376–388. doi: 10.1016/j.molcel.2014.12.004. - DOI - PMC - PubMed
-
- Trappl K, Polacek N. The ribosome: a molecular machine powered by RNA. Met Ions Life Sci. 2011;9:253–275. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
