

Optimized Mass Spectrometry Detection of Thyroid Hormones and Polar Metabolites in Rodent Cerebrospinal Fluid
- PMID: 38392972
- PMCID: PMC10890085
- DOI: 10.3390/metabo14020079
Optimized Mass Spectrometry Detection of Thyroid Hormones and Polar Metabolites in Rodent Cerebrospinal Fluid
Abstract
Thyroid hormones (TH) are required for brain development and function. Cerebrospinal fluid (CSF), which bathes the brain and spinal cord, contains TH as free hormones or as bound to transthyretin (TTR). Tight TH level regulation in the central nervous system is essential for developmental gene expression, which governs neurogenesis, myelination, and synaptogenesis. This integrated function of TH highlights the importance of developing precise and reliable methods for assessing TH levels in CSF. We report an optimized liquid chromatography-mass spectrometry (LC-MS)-based method to measure TH in rodent CSF and serum, applicable to both fresh and frozen samples. Using this new method, we find distinct differences in CSF TH in pregnant dams vs. non-pregnant adults and in embryonic vs. adult CSF. Further, targeted LC-MS metabolic profiling uncovers distinct central carbon metabolism in the CSF of these populations. TH detection and metabolite profiling of related metabolic pathways open new avenues of rigorous research into CSF TH and will inform future studies on metabolic alterations in CSF during normal development.
Keywords: cerebrospinal fluid; development; mass spectrometry method; metabolomics; reverse-phase chromatography; rodent; thyroid hormone.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
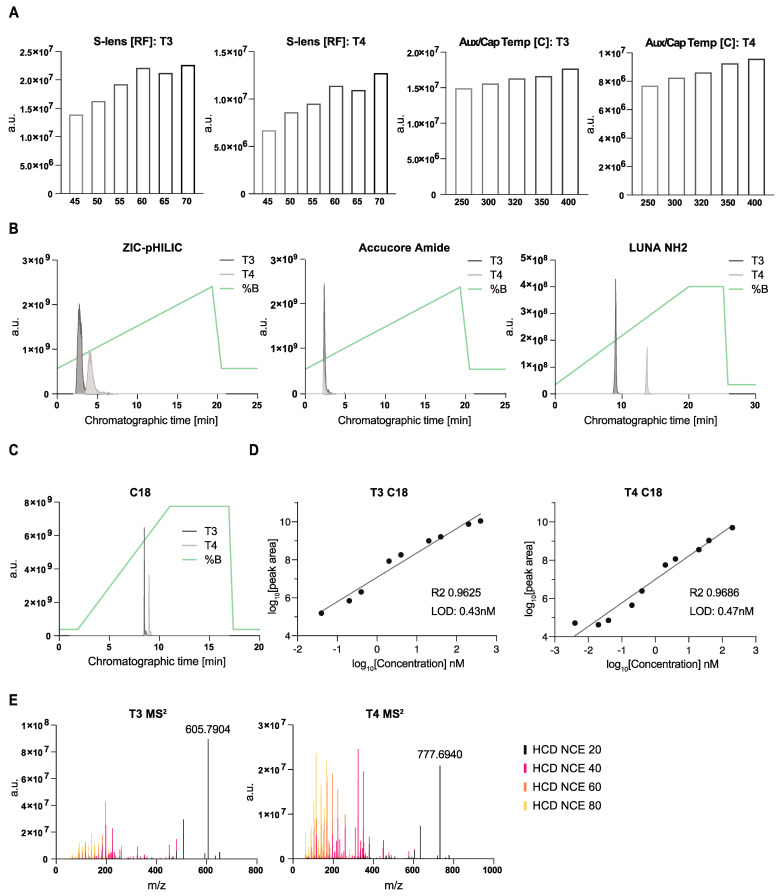
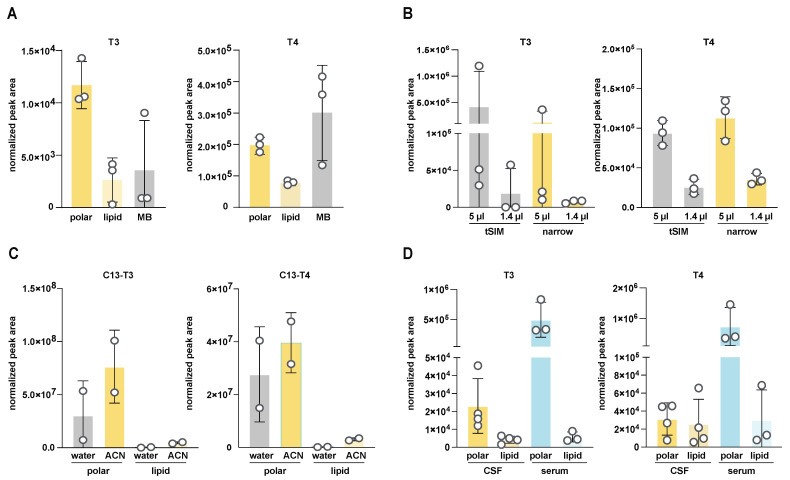
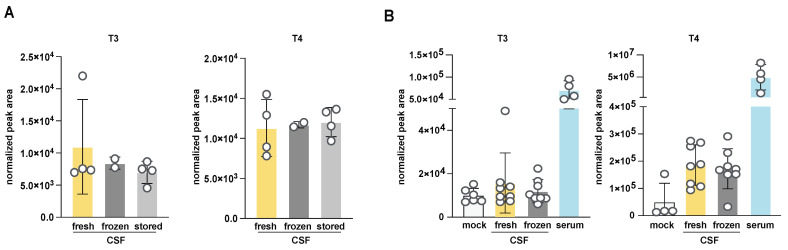
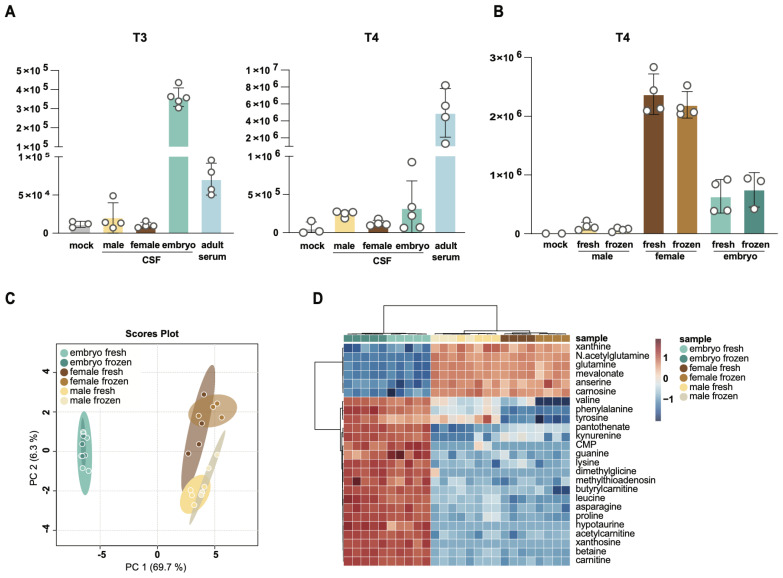
Update of
-
Optimized Mass Spectrometry Detection of Thyroid Hormones and Polar Metabolites in Rodent Cerebrospinal Fluid.bioRxiv [Preprint]. 2023 Dec 8:2023.12.07.570731. doi: 10.1101/2023.12.07.570731. bioRxiv. 2023. Update in: Metabolites. 2024 Jan 23;14(2):79. doi: 10.3390/metabo14020079. PMID: 38116027 Free PMC article. Updated. Preprint.
Similar articles
-
Optimized Mass Spectrometry Detection of Thyroid Hormones and Polar Metabolites in Rodent Cerebrospinal Fluid.bioRxiv [Preprint]. 2023 Dec 8:2023.12.07.570731. doi: 10.1101/2023.12.07.570731. bioRxiv. 2023. Update in: Metabolites. 2024 Jan 23;14(2):79. doi: 10.3390/metabo14020079. PMID: 38116027 Free PMC article. Updated. Preprint.
-
Expression of thyroid hormone transporters and deiodinases at the brain barriers in the embryonic chicken: Insights into the regulation of thyroid hormone availability during neurodevelopment.Gen Comp Endocrinol. 2015 Apr 1;214:30-9. doi: 10.1016/j.ygcen.2015.02.021. Epub 2015 Mar 5. Gen Comp Endocrinol. 2015. PMID: 25745816
-
Regulators of thyroid hormone availability and action in embryonic chicken brain development.Gen Comp Endocrinol. 2013 Sep 1;190:96-104. doi: 10.1016/j.ygcen.2013.05.003. Epub 2013 May 21. Gen Comp Endocrinol. 2013. PMID: 23707378 Review.
-
Exploring the aging process of cognitively healthy adults by analyzing cerebrospinal fluid metabolomics using liquid chromatography-tandem mass spectrometry.BMC Geriatr. 2023 Apr 5;23(1):217. doi: 10.1186/s12877-023-03939-6. BMC Geriatr. 2023. PMID: 37020298 Free PMC article.
-
Thyroid Hormone Distributor Proteins During Development in Vertebrates.Front Endocrinol (Lausanne). 2019 Aug 8;10:506. doi: 10.3389/fendo.2019.00506. eCollection 2019. Front Endocrinol (Lausanne). 2019. PMID: 31440205 Free PMC article. Review.
Cited by
-
Brain development and bioenergetic changes.Neurobiol Dis. 2024 Sep;199:106550. doi: 10.1016/j.nbd.2024.106550. Epub 2024 Jun 6. Neurobiol Dis. 2024. PMID: 38849103 Free PMC article. Review.
-
Harnessing the circadian nature of the choroid plexus and cerebrospinal fluid.NPJ Biol Timing Sleep. 2025;2(1):19. doi: 10.1038/s44323-025-00033-5. Epub 2025 May 26. NPJ Biol Timing Sleep. 2025. PMID: 40438730 Free PMC article. Review.
References
-
- Roberts L.M., Woodford K., Zhou M., Black D.S., Haggerty J.E., Tate E.H., Grindstaff K.K., Mengesha W., Raman C., Zerangue N. Expression of the Thyroid Hormone Transporters Monocarboxylate Transporter-8 (SLC16A2) and Organic Ion Transporter-14 (SLCO1C1) at the Blood-Brain Barrier. Endocrinology. 2008;149:6251–6261. doi: 10.1210/en.2008-0378. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
