

IL-27 regulates the differentiation of follicular helper NKT cells via metabolic adaptation of mitochondria
- PMID: 38394242
- PMCID: PMC10907256
- DOI: 10.1073/pnas.2313964121
IL-27 regulates the differentiation of follicular helper NKT cells via metabolic adaptation of mitochondria
Abstract
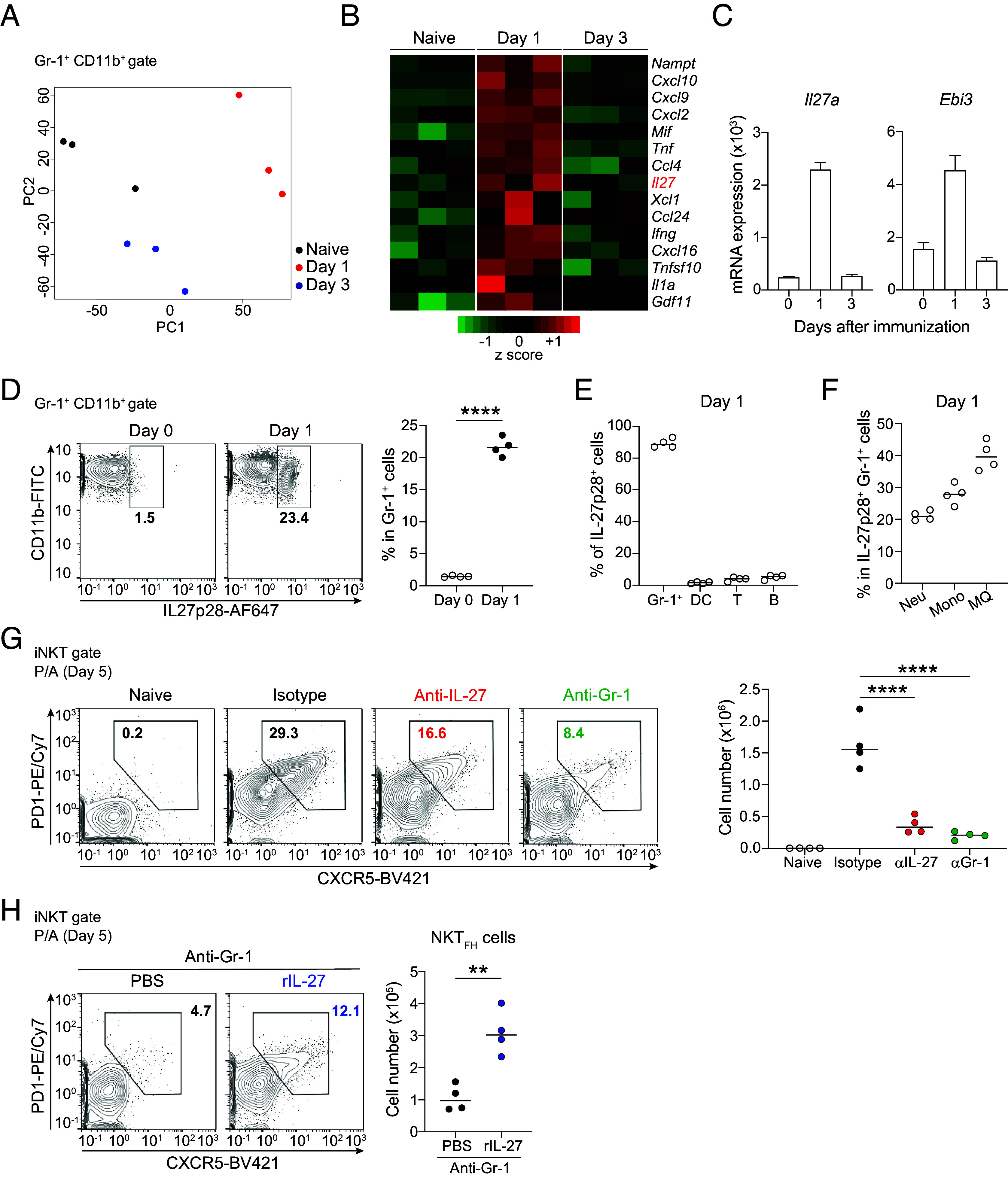
Invariant natural killer T (iNKT) cells are innate-like T lymphocytes that express an invariant T cell receptor α chain and contribute to bridging innate and acquired immunity with rapid production of large amounts of cytokines after stimulation. Among effecter subsets of iNKT cells, follicular helper NKT (NKTFH) cells are specialized to help B cells. However, the mechanisms of NKTFH cell differentiation remain to be elucidated. In this report, we studied the mechanism of NKTFH cell differentiation induced by pneumococcal surface protein A and α-galactosylceramide (P/A) vaccination. We found that Gr-1+ cells helped iNKT cell proliferation and NKTFH cell differentiation in the spleen by producing interleukin-27 (IL-27) in the early phase after vaccination. The neutralization of IL-27 impaired NKTFH cell differentiation, which resulted in compromised antibody production and diminished protection against Streptococcus pneumoniae infection by the P/A vaccine. Our data indicated that Gr-1+ cell-derived IL-27 stimulated mitochondrial metabolism, meeting the energic demand required for iNKT cells to differentiate into NKTFH cells. Interestingly, Gr-1+ cell-derived IL-27 was induced by iNKT cells via interferon-γ production. Collectively, our findings suggest that optimizing the metabolism of iNKT cells was essential for acquiring specific effector functions, and they provide beneficial knowledge on iNKT cell-mediated vaccination-mediated therapeutic strategies.
Keywords: IL-27; Streptococcus pneumoniae; follicular helper; iNKT; mitochondrial metabolism.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures




References
-
- Brennan P. J., Brigl M., Brenner M. B., Invariant natural killer T cells: An innate activation scheme linked to diverse effector functions. Nat. Rev. Immunol. 13, 101–117 (2013). - PubMed
-
- Brigl M., Brenner M. B., CD1: Antigen presentation and T cell function. Annu. Rev. Immunol. 22, 817–890 (2004). - PubMed
-
- Kronenberg M., Toward an understanding of NKT cell biology: Progress and paradoxes. Annu. Rev. Immunol. 23, 877–900 (2005). - PubMed
-
- Taniguchi M., Harada M., Kojo S., Nakayama T., Wakao H., The regulatory role of Valpha14 NKT cells in innate and acquired immune response. Annu. Rev. Immunol. 21, 483–513 (2003). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
