

Exploring the Micro-Mosaic Landscape of FGFR3 Mutations in the Ageing Male Germline and Their Potential Implications in Meiotic Differentiation
- PMID: 38397181
- PMCID: PMC10888257
- DOI: 10.3390/genes15020191
Exploring the Micro-Mosaic Landscape of FGFR3 Mutations in the Ageing Male Germline and Their Potential Implications in Meiotic Differentiation
Abstract
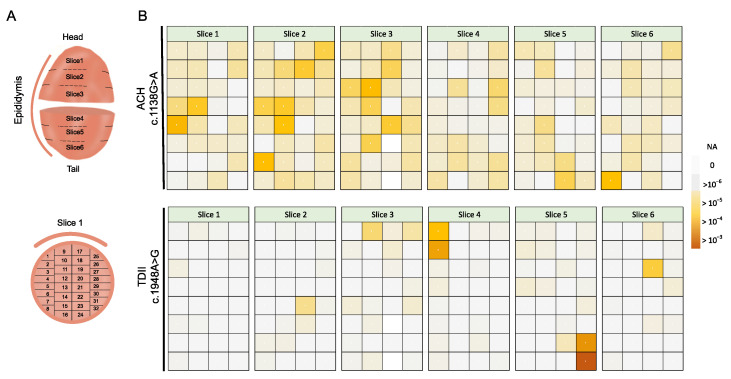
Advanced paternal age increases the risk of transmitting de novo germline mutations, particularly missense mutations activating the receptor tyrosine kinase (RTK) signalling pathway, as exemplified by the FGFR3 mutation, which is linked to achondroplasia (ACH). This risk is attributed to the expansion of spermatogonial stem cells carrying the mutation, forming sub-clonal clusters in the ageing testis, thereby increasing the frequency of mutant sperm and the number of affected offspring from older fathers. While prior studies proposed a correlation between sub-clonal cluster expansion in the testis and elevated mutant sperm production in older donors, limited data exist on the universality of this phenomenon. Our study addresses this gap by examining the testis-expansion patterns, as well as the increases in mutations in sperm for two FGFR3 variants-c.1138G>A (p.G380R) and c.1948A>G (p.K650E)-which are associated with ACH or thanatophoric dysplasia (TDII), respectively. Unlike the ACH mutation, which showed sub-clonal expansion events in an aged testis and a significant increase in mutant sperm with the donor's age, as also reported in other studies, the TDII mutation showed focal mutation pockets in the testis but exhibited reduced transmission into sperm and no significant age-related increase. The mechanism behind this divergence remains unclear, suggesting potential pleiotropic effects of aberrant RTK signalling in the male germline, possibly hindering differentiation requiring meiosis. This study provides further insights into the transmission risks of micro-mosaics associated with advanced paternal age in the male germline.
Keywords: FGFR3; achondroplasia; congenital disorders; driver mutations; germline mutagenesis; receptor tyrosine kinase; thanatophoric dysplasia.
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
Exploring FGFR3 Mutations in the Male Germline: Implications for Clonal Germline Expansions and Paternal Age-Related Dysplasias.Genome Biol Evol. 2024 Feb 1;16(2):evae015. doi: 10.1093/gbe/evae015. Genome Biol Evol. 2024. PMID: 38411226 Free PMC article.
-
Cellular evidence for selfish spermatogonial selection in aged human testes.Andrology. 2014 May;2(3):304-14. doi: 10.1111/j.2047-2927.2013.00175.x. Epub 2013 Dec 19. Andrology. 2014. PMID: 24357637 Review.
-
Severe achondroplasia due to two de novo variants in the transmembrane domain of FGFR3 on the same allele: A case report.Mol Genet Genomic Med. 2020 Mar;8(3):e1148. doi: 10.1002/mgg3.1148. Epub 2020 Jan 23. Mol Genet Genomic Med. 2020. PMID: 31975530 Free PMC article.
-
Graded activation of fibroblast growth factor receptor 3 by mutations causing achondroplasia and thanatophoric dysplasia.Nat Genet. 1996 Jun;13(2):233-7. doi: 10.1038/ng0696-233. Nat Genet. 1996. PMID: 8640234
-
Paternal age effect mutations and selfish spermatogonial selection: causes and consequences for human disease.Am J Hum Genet. 2012 Feb 10;90(2):175-200. doi: 10.1016/j.ajhg.2011.12.017. Am J Hum Genet. 2012. PMID: 22325359 Free PMC article. Review.
Cited by
-
Exploring FGFR3 Mutations in the Male Germline: Implications for Clonal Germline Expansions and Paternal Age-Related Dysplasias.Genome Biol Evol. 2024 Feb 1;16(2):evae015. doi: 10.1093/gbe/evae015. Genome Biol Evol. 2024. PMID: 38411226 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
