

One-line hybrid rice with high-efficiency synthetic apomixis and near-normal fertility
- PMID: 38400858
- PMCID: PMC10894110
- DOI: 10.1007/s00299-024-03154-6
One-line hybrid rice with high-efficiency synthetic apomixis and near-normal fertility
Abstract
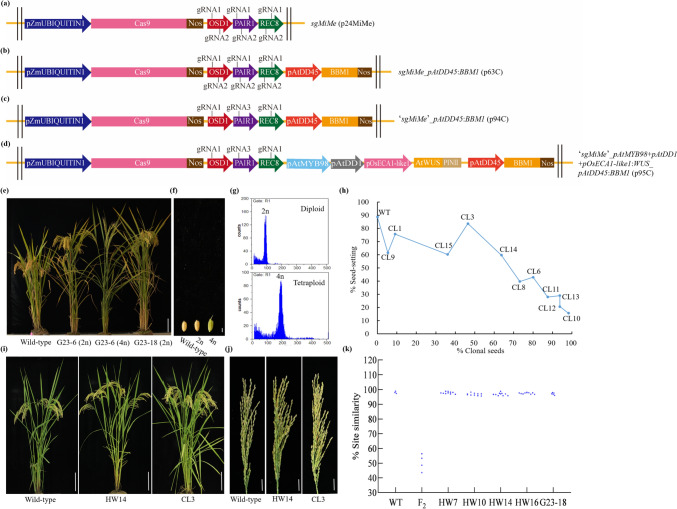
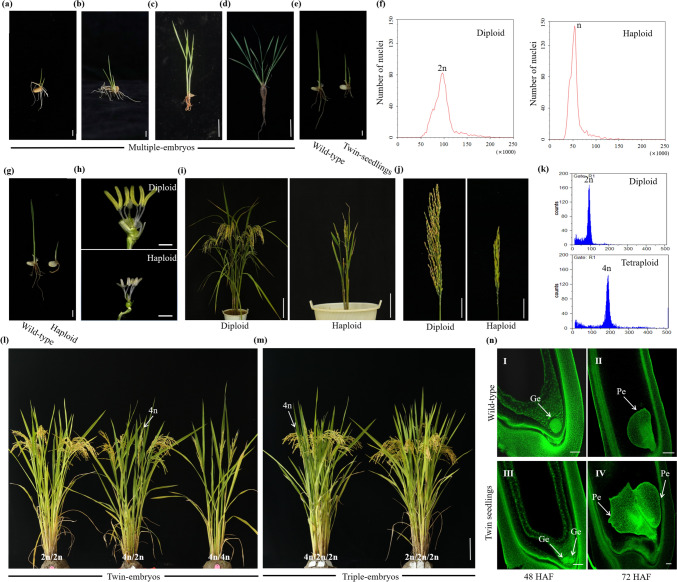
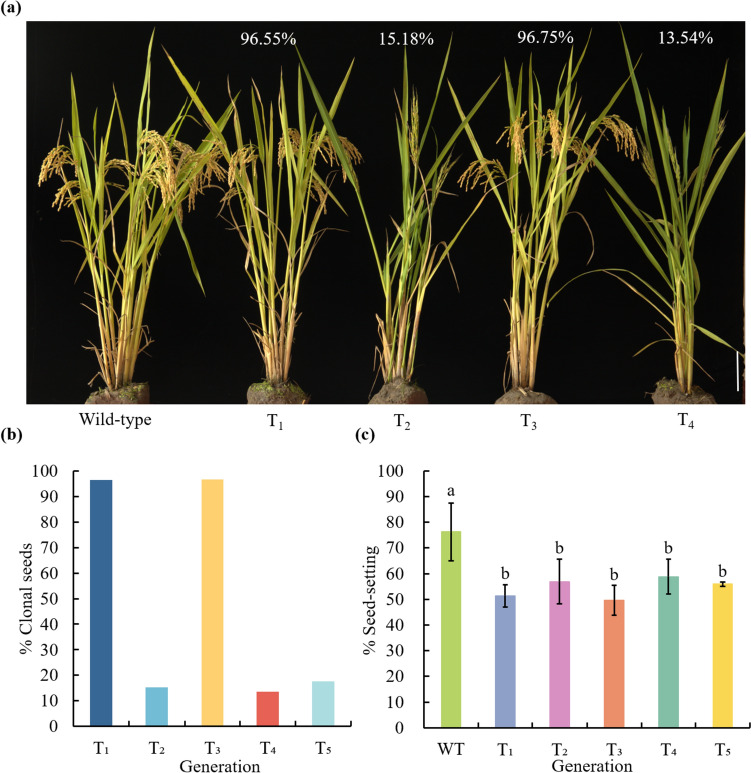
High-frequency clonal seeds and near-normal fertility were obtained by engineering synthetic apomixis in hybrid rice. The one-line strategy, with the advantage of unnecessary seed production, is the final stage for the hybrid rice development and can be achieved through the fixation of heterosis via artificially inducing apomixis. Recently, synthetic apomixis has been generated in rice by combining MiMe (Mitosis instead of Meiosis) with either the ectopic expression of BABY BOOM (BBM1 or BBM4) or mutation of MATRILINEAL (MTL), resulting in over 95.00% of clonal seeds. However, the frequency of clonal seeds was only 29.20% when AtDD45 promoter was used to drive BBM1. In addition, achieving both a high frequency of clonal seeds and near-normal fertility simultaneously had been elusive in earlier strategies. In this study, using AtDD45 promoter to drive BBM1 expression in combination with the MiMe mutant resulted in the apomixis frequency as high as 98.70%. Even more, employing fusion promoters (AtMYB98_AtDD1_OsECA1-like1) to drive WUS expression in combination with pAtDD45:BBM1 and MiMe could produce clonal seeds at rates of up to 98.21%, the highest seed setting rate reached to 83.67%. Multiple-embryos were observed in clonal lines at a frequency ranging from 3.37% to 60.99%. Transmission of the high frequency of apomixis through skipped generations (atavism) was identified in two clonal lines, even though it remained stable in the majority of clonal lines. These findings significantly advance the pursuit of fixed heterosis in rice through synthetic apomixis, edging closer to its agricultural application.
Keywords: Apomixis; Atavism; Multiple-embryos; One-line hybrid rice.
© 2024. The Author(s).
Conflict of interest statement
The authors have no competing interests to declare that are relevant to the content of this article.
Figures
References
-
- Cao ML, Xia YM, Zhan YJ et al (2019) A seed sorting method for fixed plant heterosis. CN201910307717.3
-
- Doyle JJT, Doyle JL. Isolation of DNA from fresh tissue. Focus. 1990;12:13–15.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
