

5-Formylcytosine mediated DNA-peptide cross-link induces predominantly semi-targeted mutations in both Escherichia coli and human cells
- PMID: 38401843
- PMCID: PMC10966706
- DOI: 10.1016/j.jbc.2024.105786
5-Formylcytosine mediated DNA-peptide cross-link induces predominantly semi-targeted mutations in both Escherichia coli and human cells
Abstract
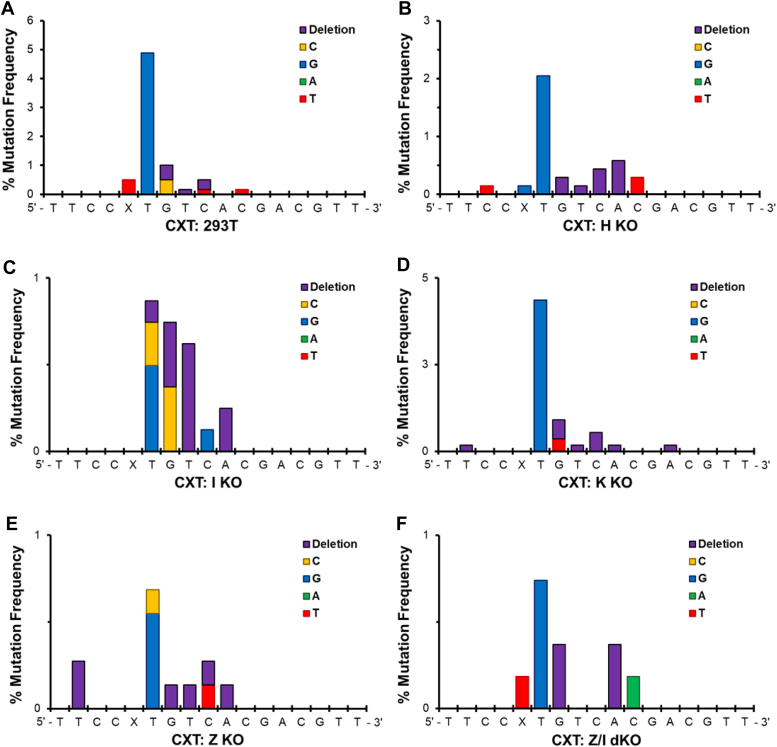
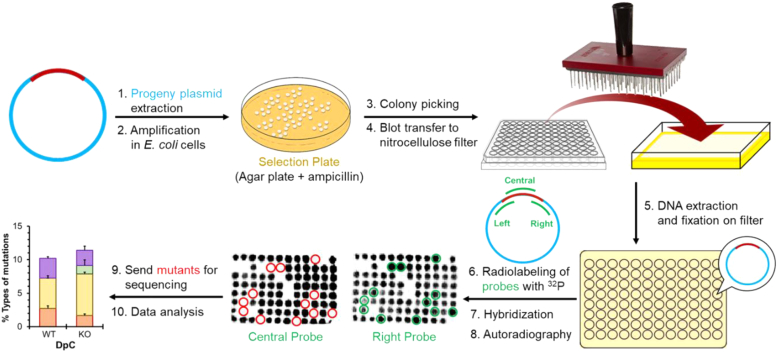
Histone proteins can become trapped on DNA in the presence of 5-formylcytosine (5fC) to form toxic DNA-protein conjugates. Their repair may involve proteolytic digestion resulting in DNA-peptide cross-links (DpCs). Here, we have investigated replication of a model DpC comprised of an 11-mer peptide (NH2-GGGKGLGK∗GGA) containing an oxy-lysine residue (K∗) conjugated to 5fC in DNA. Both CXG and CXT (where X = 5fC-DpC) sequence contexts were examined. Replication of both constructs gave low viability (<10%) in Escherichia coli, whereas TLS efficiency was high (72%) in HEK 293T cells. In E. coli, the DpC was bypassed largely error-free, inducing only 2 to 3% mutations, which increased to 4 to 5% with SOS. For both sequences, semi-targeted mutations were dominant, and for CXG, the predominant mutations were G→T and G→C at the 3'-base to the 5fC-DpC. In HEK 293T cells, 7 to 9% mutations occurred, and the dominant mutations were the semi-targeted G → T for CXG and T → G for CXT. These mutations were reduced drastically in cells deficient in hPol η, hPol ι or hPol ζ, suggesting a role of these TLS polymerases in mutagenic TLS. Steady-state kinetics studies using hPol η confirmed that this polymerase induces G → T and T → G transversions at the base immediately 3' to the DpC. This study reveals a unique replication pattern of 5fC-conjugated DpCs, which are bypassed largely error-free in both E. coli and human cells and induce mostly semi-targeted mutations at the 3' position to the lesion.
Keywords: 5-formylcytosine; DNA polymerase; DNA replication; DNA-protein crosslinks; mutagenesis; peptides; protein-DNA interaction; translesion synthesis.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare no conflicts of interest with the contents of this article.
Figures







References
-
- Barker S., Weinfeld M., Murray D. DNA-protein crosslinks: their induction, repair, and biological consequences. Mutat. Res. 2005;589:111–135. - PubMed
-
- Basu A.K., Campbell C., Tretyakova N.Y. In: DNA Damage, DNA Repair and Disease. Dizdaroglu M.a.L., R S., editors. Royal Society of Chemistry; London, UK: 2021. DNA-protein cross-links: formation, genotoxicity and repair; pp. 154–174.
-
- Ide H., Shoulkamy M.I., Nakano T., Miyamoto-Matsubara M., Salem A.M. Repair and biochemical effects of DNA-protein crosslinks. Mutat. Res. 2011;711:113–122. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
