

NAAA-regulated lipid signaling in monocytes controls the induction of hyperalgesic priming in mice
- PMID: 38402219
- PMCID: PMC10894261
- DOI: 10.1038/s41467-024-46139-5
NAAA-regulated lipid signaling in monocytes controls the induction of hyperalgesic priming in mice
Abstract
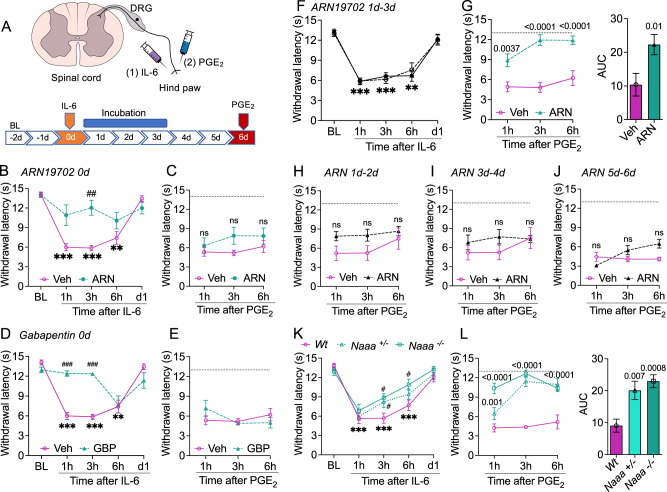
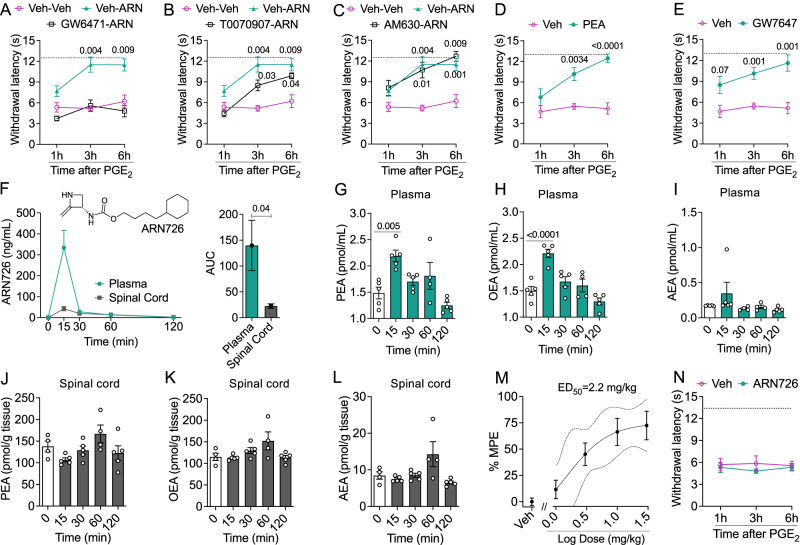
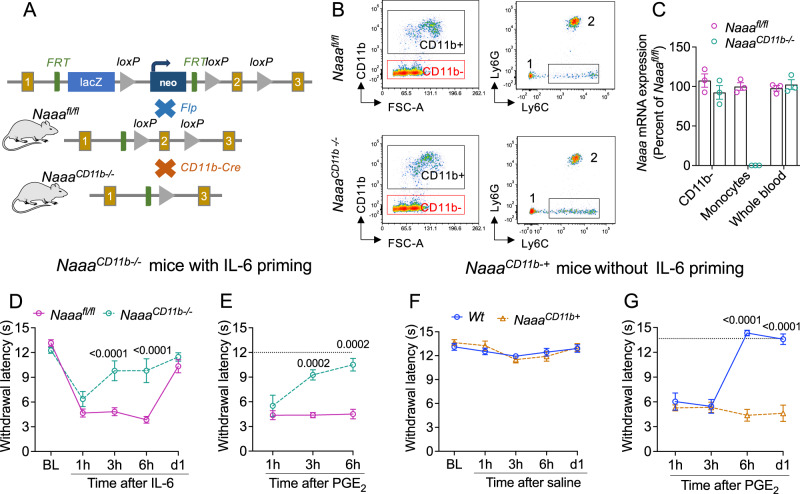
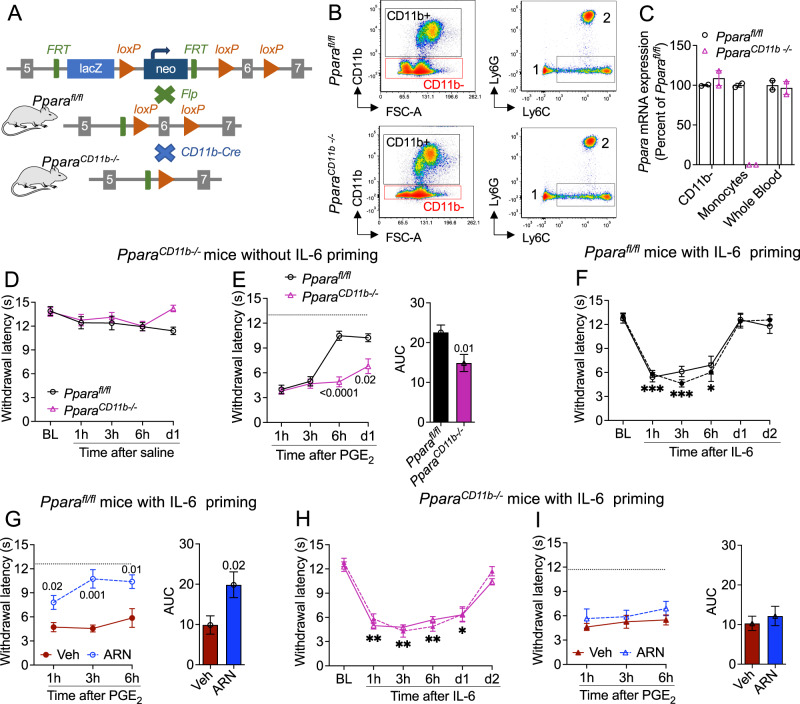
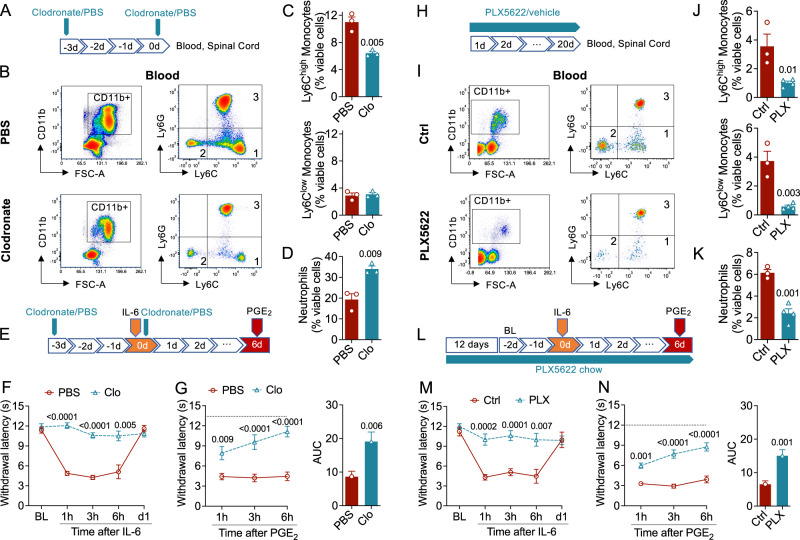
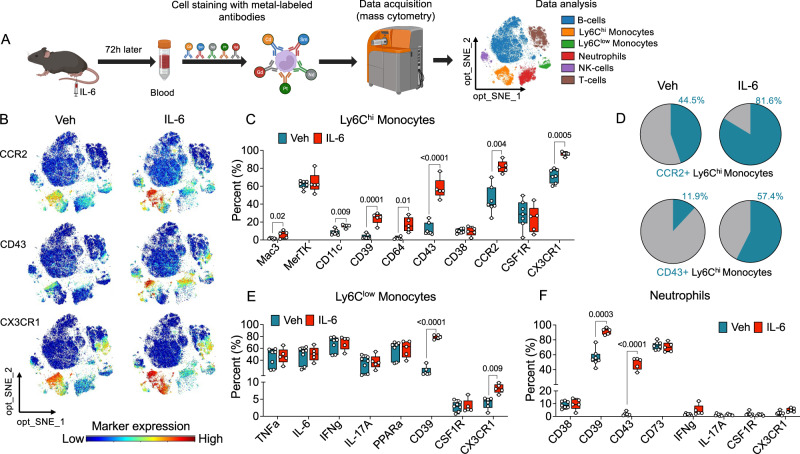
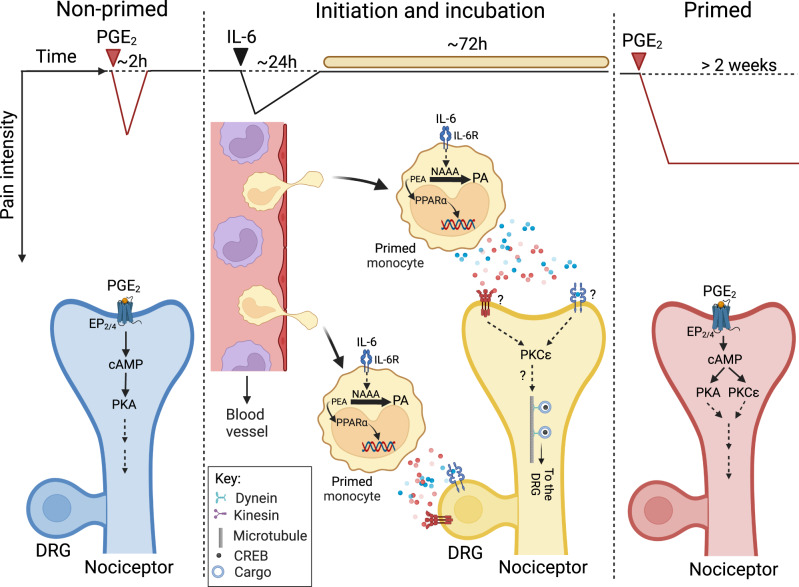
Circulating monocytes participate in pain chronification but the molecular events that cause their deployment are unclear. Using a mouse model of hyperalgesic priming (HP), we show that monocytes enable progression to pain chronicity through a mechanism that requires transient activation of the hydrolase, N-acylethanolamine acid amidase (NAAA), and the consequent suppression of NAAA-regulated lipid signaling at peroxisome proliferator-activated receptor-α (PPAR-α). Inhibiting NAAA in the 72 hours following administration of a priming stimulus prevented HP. This effect was phenocopied by NAAA deletion and depended on PPAR-α recruitment. Mice lacking NAAA in CD11b+ cells - monocytes, macrophages, and neutrophils - were resistant to HP induction. Conversely, mice overexpressing NAAA or lacking PPAR-α in the same cells were constitutively primed. Depletion of monocytes, but not resident macrophages, generated mice that were refractory to HP. The results identify NAAA-regulated signaling in monocytes as a control node in the induction of HP and, potentially, the transition to pain chronicity.
© 2024. The Author(s).
Conflict of interest statement
D.P. and M.M. are inventors in patents that protect ARN19702 and other NAAA inhibitors, owned by the University of California, the University of Parma, the University of Urbino, and the Fondazione Istituto Italiano di Tecnologia (no. 13/898,225, filed 20 May 2013, published 3 April 2014; no. 62/337,744, filed 17 May 2016, published 23 November 2017). D.P. and Y.F. are inventors in a patent application that protects the algostatic effects of NAAA inhibitors, filed by the University of California (no. 63/166,134, filed 25 March 2021, published 29 September 2022). The other authors declare no competing interest. No specific aspect of the manuscript was covered in patent applications.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
