

Alternative mRNA splicing events and regulators in epidermal differentiation
- PMID: 38402585
- PMCID: PMC11293371
- DOI: 10.1016/j.celrep.2024.113814
Alternative mRNA splicing events and regulators in epidermal differentiation
Abstract
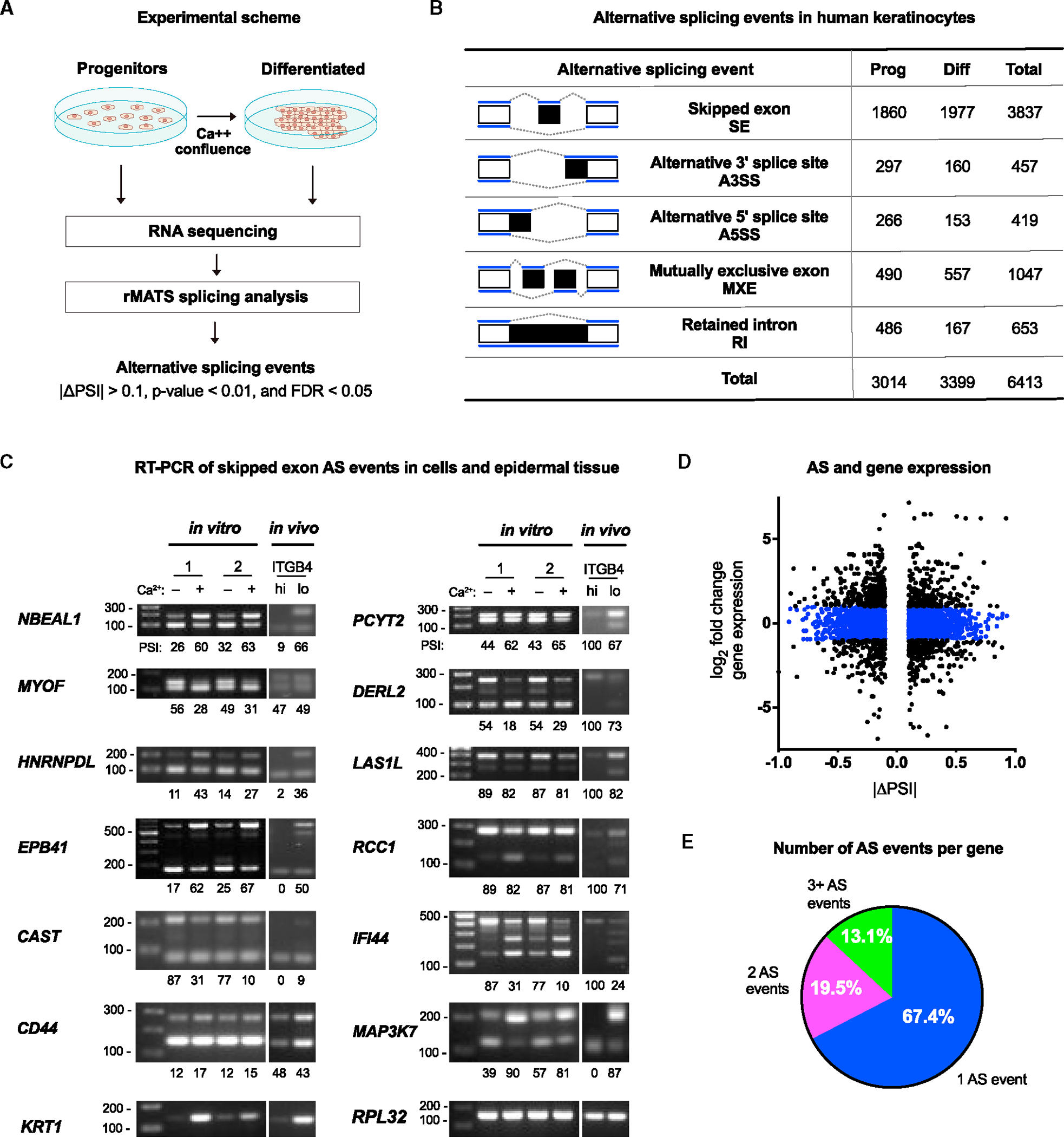
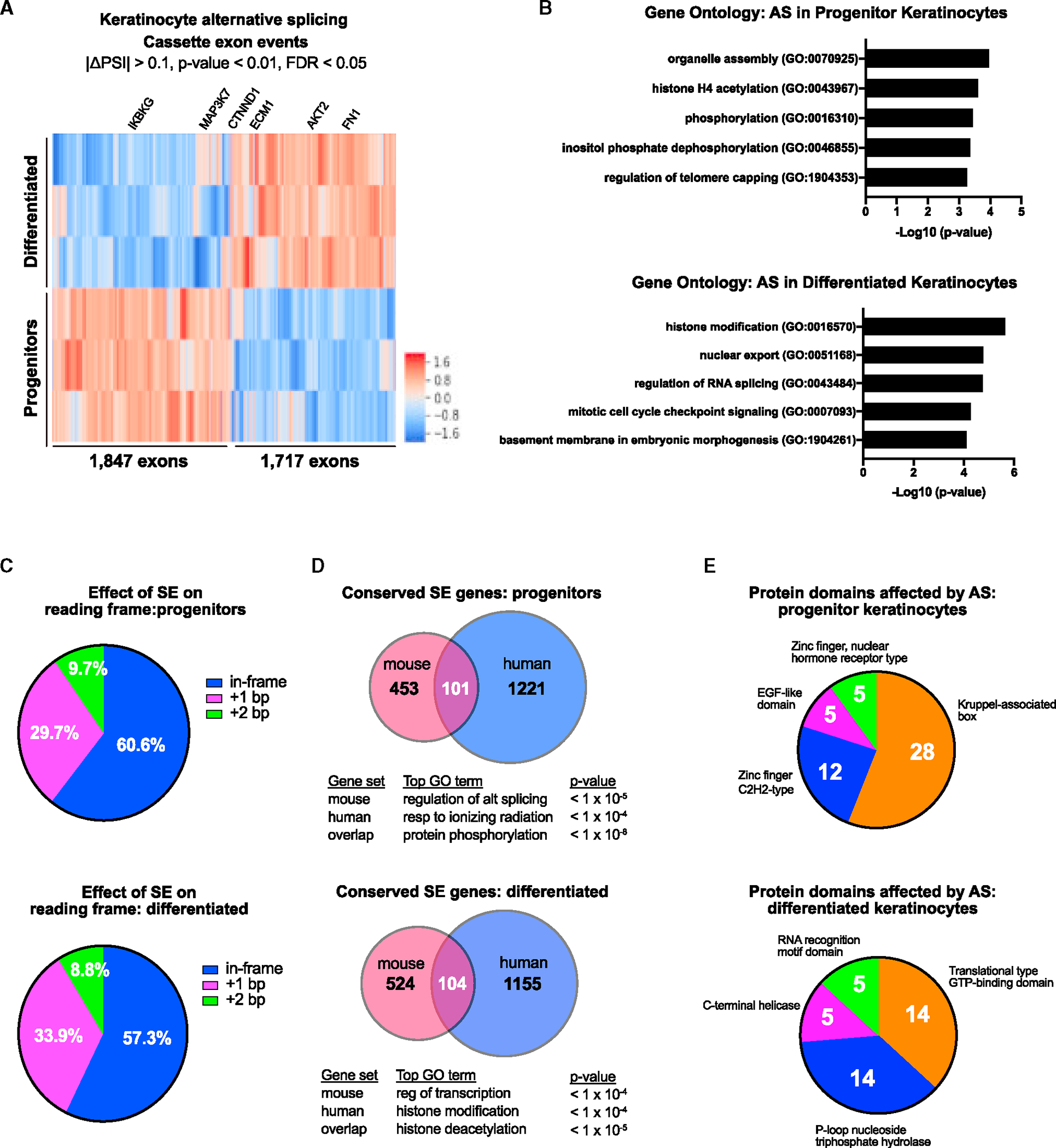
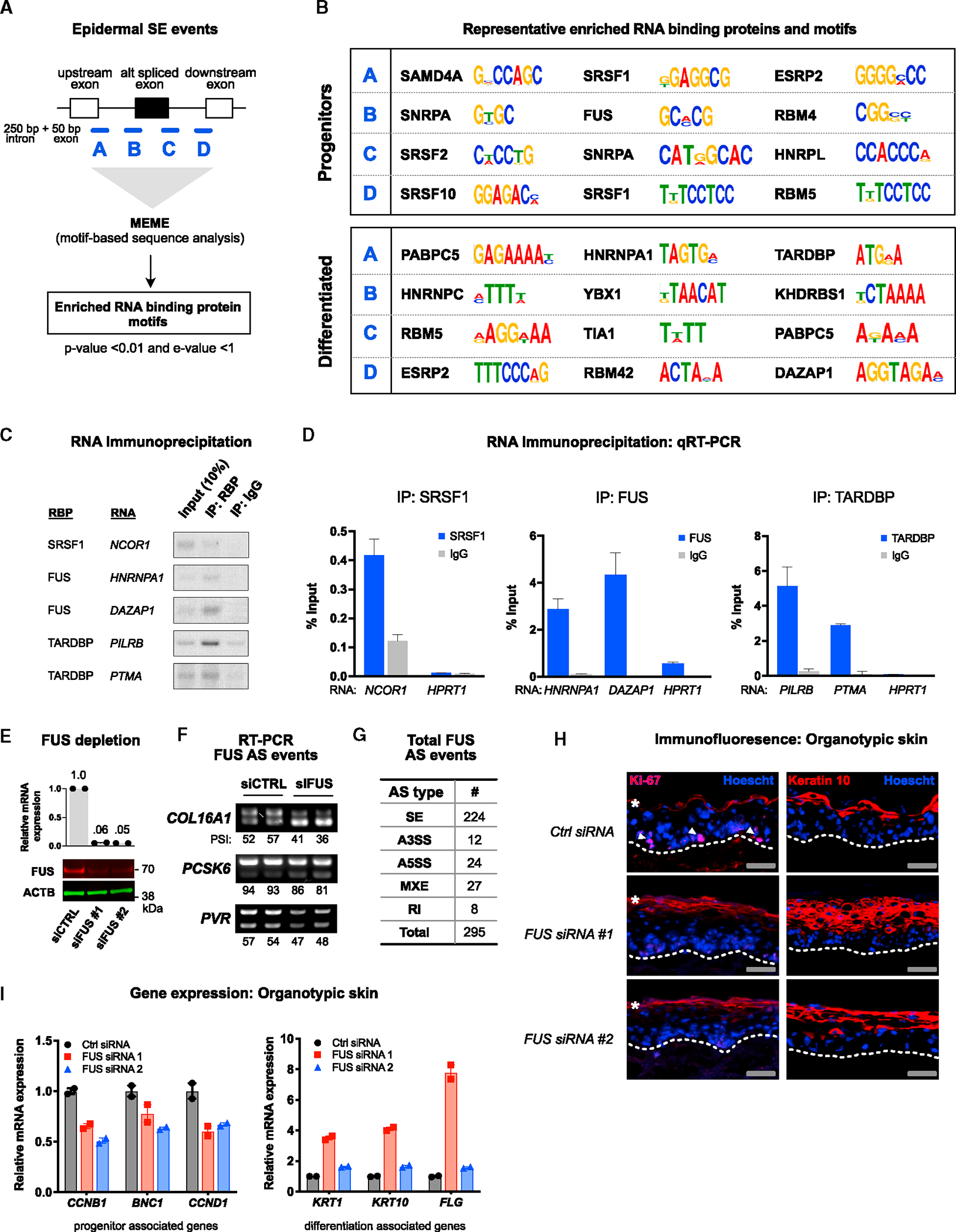
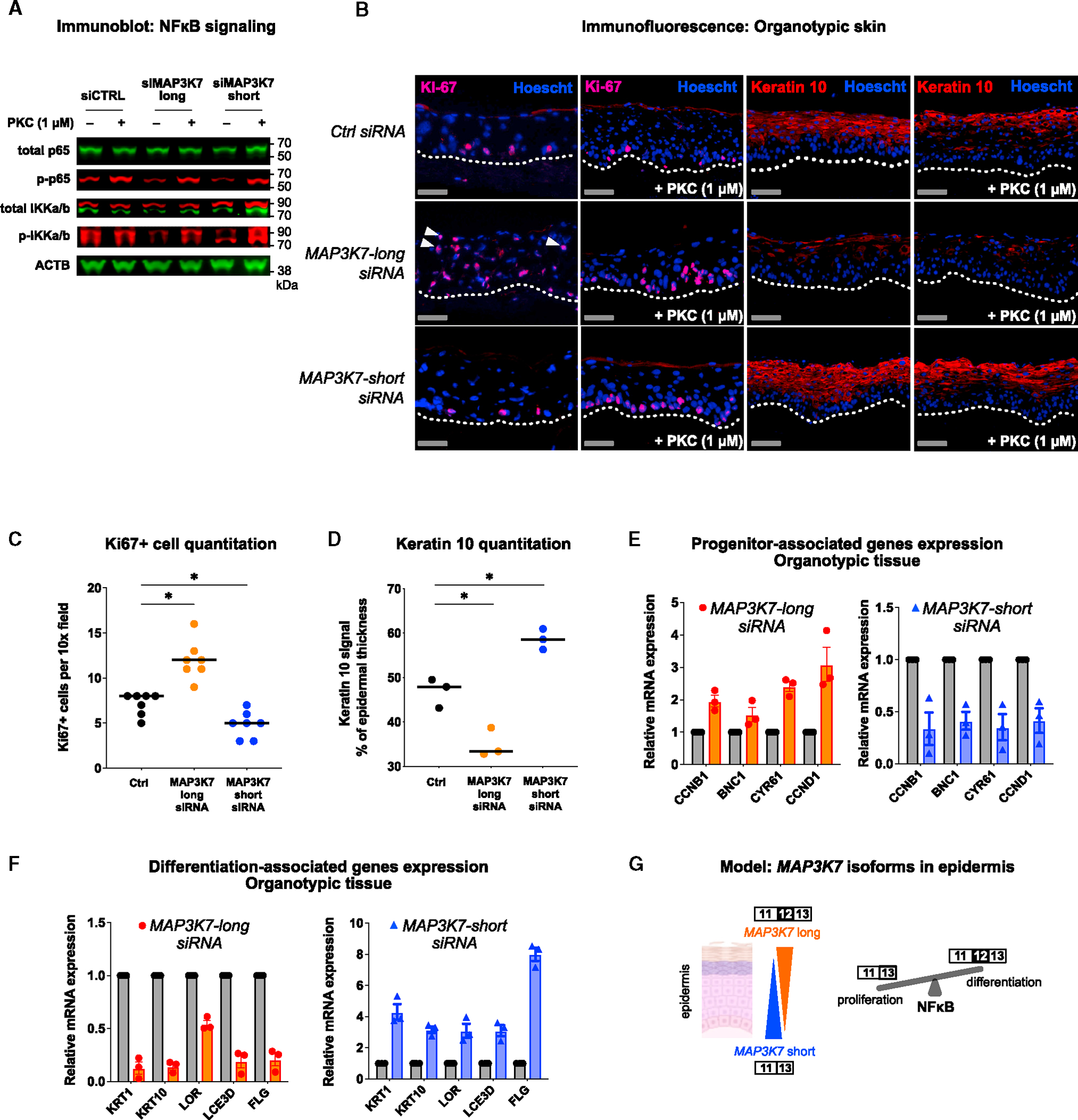
Alternative splicing (AS) of messenger RNAs occurs in ∼95% of multi-exon human genes and generates diverse RNA and protein isoforms. We investigated AS events associated with human epidermal differentiation, a process crucial for skin function. We identified 6,413 AS events, primarily involving cassette exons. We also predicted 34 RNA-binding proteins (RBPs) regulating epidermal AS, including 19 previously undescribed candidate regulators. From these results, we identified FUS as an RBP that regulates the balance between keratinocyte proliferation and differentiation. Additionally, we characterized the function of a cassette exon AS event in MAP3K7, which encodes a kinase involved in cell signaling. We found that a switch from the short to long isoform of MAP3K7, triggered during differentiation, enforces the demarcation between proliferating basal progenitors and overlying differentiated strata. Our findings indicate that AS occurs extensively in the human epidermis and has critical roles in skin homeostasis.
Keywords: CP: Molecular biology; CP: Stem cell research; MAP3K7; alternative splicing; epidermal differentiation; skin; skin homeostasis.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures

Similar articles
-
From transcriptomic to protein level changes in TDP-43 and FUS loss-of-function cell models.Biochim Biophys Acta. 2015 Dec;1849(12):1398-410. doi: 10.1016/j.bbagrm.2015.10.015. Epub 2015 Oct 26. Biochim Biophys Acta. 2015. PMID: 26514432
-
OncoSplicing 3.0: an updated database for identifying RBPs regulating alternative splicing events in cancers.Nucleic Acids Res. 2025 Jan 6;53(D1):D1460-D1466. doi: 10.1093/nar/gkae1098. Nucleic Acids Res. 2025. PMID: 39558172 Free PMC article.
-
A posttranscriptional mechanism that controls Ptbp1 abundance in the Xenopus epidermis.Mol Cell Biol. 2015 Feb;35(4):758-68. doi: 10.1128/MCB.01040-14. Epub 2014 Dec 15. Mol Cell Biol. 2015. PMID: 25512611 Free PMC article.
-
Function of alternative splicing.Gene. 2013 Feb 1;514(1):1-30. doi: 10.1016/j.gene.2012.07.083. Epub 2012 Aug 15. Gene. 2013. PMID: 22909801 Free PMC article. Review.
-
Regulation of human cathepsin B by alternative mRNA splicing: homeostasis, fatal errors and cell death.Biol Chem. 2006 Aug;387(8):1017-21. doi: 10.1515/BC.2006.125. Biol Chem. 2006. PMID: 16895470 Review.
Cited by
-
FUS-Mediated Inhibition of Myogenesis Elicited by Suppressing TNNT1 Production.Mol Cell Biol. 2024;44(9):391-409. doi: 10.1080/10985549.2024.2383296. Epub 2024 Aug 12. Mol Cell Biol. 2024. PMID: 39133076 Free PMC article.
-
Splicing to orchestrate cell fate.Mol Ther Nucleic Acids. 2024 Dec 6;36(1):102416. doi: 10.1016/j.omtn.2024.102416. eCollection 2025 Mar 11. Mol Ther Nucleic Acids. 2024. PMID: 39811494 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
