

Critical roles of nicotinic acetylcholine receptors in olfactory memory formation and retrieval in crickets
- PMID: 38405118
- PMCID: PMC10884312
- DOI: 10.3389/fphys.2024.1345397
Critical roles of nicotinic acetylcholine receptors in olfactory memory formation and retrieval in crickets
Abstract
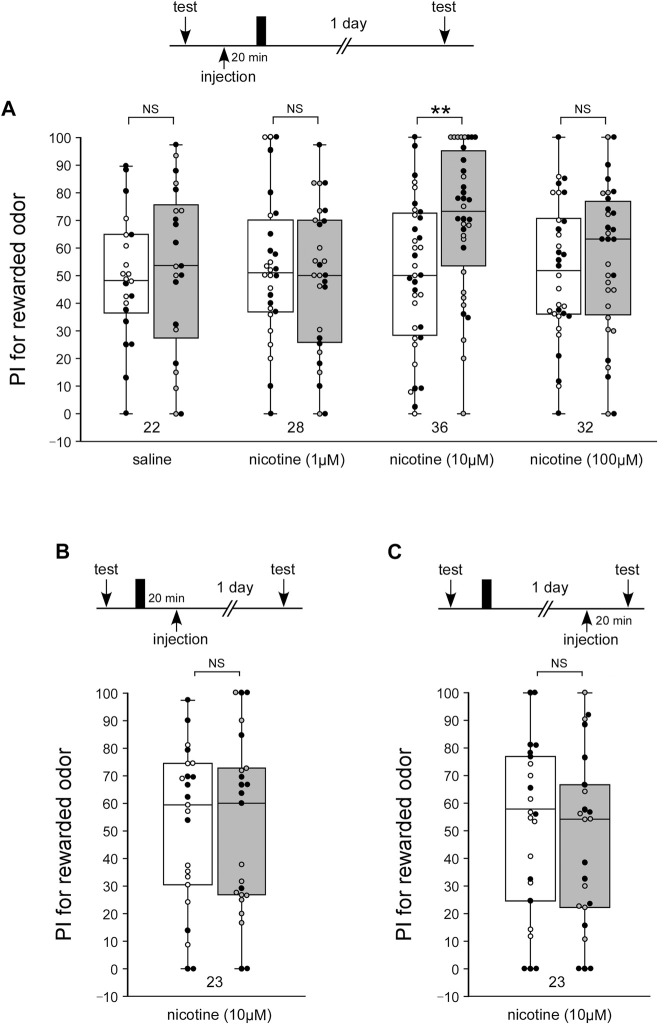
Acetylcholine (ACh) is a major excitatory neurotransmitter in the insect central nervous system, and insect neurons express several types of ACh receptors (AChRs). AChRs are classified into two subgroups, muscarinic AChRs and nicotinic AChRs (nAChRs). nAChRs are also divided into two subgroups by sensitivity to α-bungarotoxin (α-BGT). The cricket Gryllus bimaculatus is one of the useful insects for studying the molecular mechanisms in olfactory learning and memory. However, the roles of nAChRs in olfactory learning and memory of the cricket are still unknown. In the present study, to investigate whether nAChRs are involved in cricket olfactory learning and memory, we tested the effects of two different AChR antagonists on long-term memory (LTM) formation and retrieval in a behavioral assay. The two AChR antagonists that we used are mecamylamine (MEC), an α-BGT-insensitive nAChR antagonist, and methyllycaconitine (MLA), an α-BGT-sensitive nAChR antagonist. In crickets, multiple-trial olfactory conditioning induced 1-day memory (LTM), whereas single-trial olfactory conditioning induced 1-h memory (mid-term memory, MTM) but not 1-day memory. Crickets injected with MEC 20 min before the retention test at 1 day after the multiple-trial conditioning exhibited no memory retrieval. This indicates that α-BGT-insensitive nAChRs participate in memory retrieval. In addition, crickets injected with MLA before the multiple-trial conditioning exhibited MTM but not LTM, indicating that α-BGT-sensitive nAChRs participate in the formation of LTM. Moreover, injection of nicotine (an nAChR agonist) before the single-trial conditioning induced LTM. Finally, the nitric oxide (NO)-cGMP signaling pathway is known to participate in the formation of LTM in crickets, and we conducted co-injection experiments with an agonist or inhibitor of the nAChR and an activator or inhibitor of the NO-cGMP signaling pathway. The results suggest that nAChR works upstream of the NO-cGMP signaling system in the LTM formation process.
Keywords: cricket; long-term memory; mecamylamine; methyllycaconitine; nicotine; nicotinic acetylcholine receptors; olfactory learning.
Copyright © 2024 Matsumoto, Matsumoto and Mizunami.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures




Similar articles
-
State of the art on insect nicotinic acetylcholine receptor function in learning and memory.Adv Exp Med Biol. 2010;683:97-115. doi: 10.1007/978-1-4419-6445-8_9. Adv Exp Med Biol. 2010. PMID: 20737792 Review.
-
Involvement of alpha-bungarotoxin-sensitive nicotinic receptors in long-term memory formation in the honeybee (Apis mellifera).Neurobiol Learn Mem. 2006 Sep;86(2):164-74. doi: 10.1016/j.nlm.2006.02.003. Epub 2006 Apr 17. Neurobiol Learn Mem. 2006. PMID: 16616529
-
Activation of NO-cGMP Signaling Rescues Age-Related Memory Impairment in Crickets.Front Behav Neurosci. 2016 Aug 26;10:166. doi: 10.3389/fnbeh.2016.00166. eCollection 2016. Front Behav Neurosci. 2016. PMID: 27616985 Free PMC article.
-
Roles of calcium/calmodulin-dependent kinase II in long-term memory formation in crickets.PLoS One. 2014 Sep 12;9(9):e107442. doi: 10.1371/journal.pone.0107442. eCollection 2014. PLoS One. 2014. PMID: 25215889 Free PMC article.
-
Signaling Pathways for Long-Term Memory Formation in the Cricket.Front Psychol. 2018 Jun 22;9:1014. doi: 10.3389/fpsyg.2018.01014. eCollection 2018. Front Psychol. 2018. PMID: 29988479 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources