

This is a preprint.
DNA methylation-dependent and -independent binding of CDX2 directs activation of distinct developmental and homeostatic genes
- PMID: 38405700
- PMCID: PMC10888781
- DOI: 10.1101/2024.02.11.579850
DNA methylation-dependent and -independent binding of CDX2 directs activation of distinct developmental and homeostatic genes
Abstract
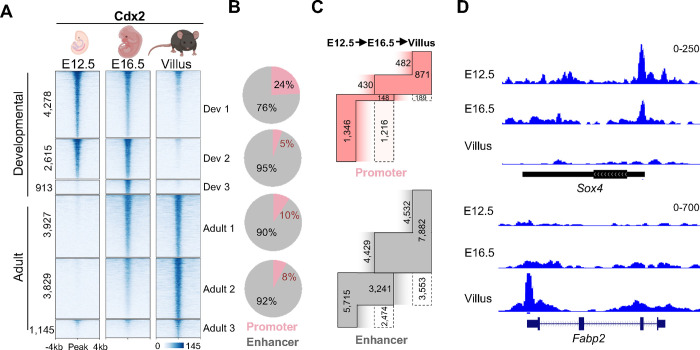
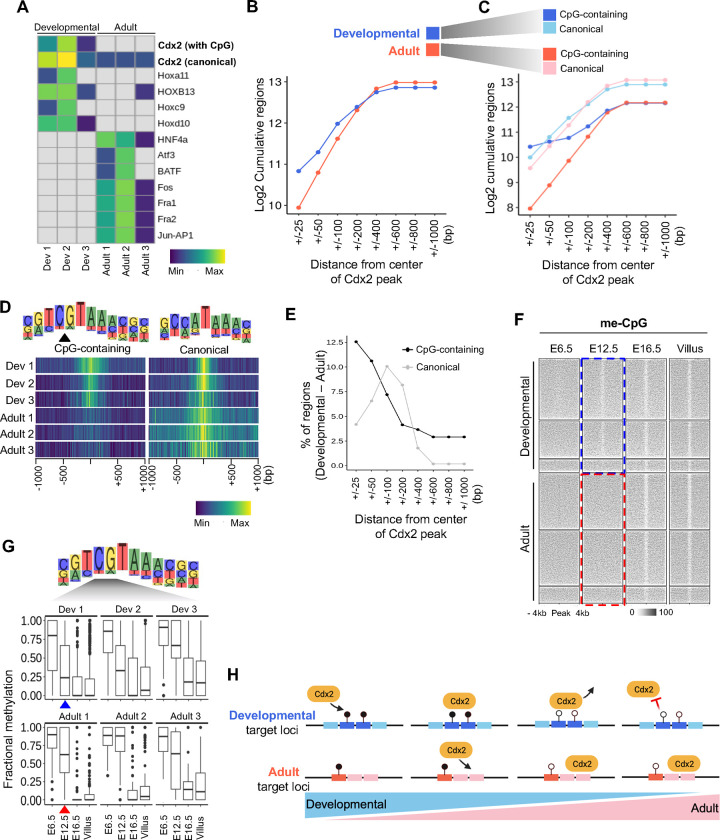
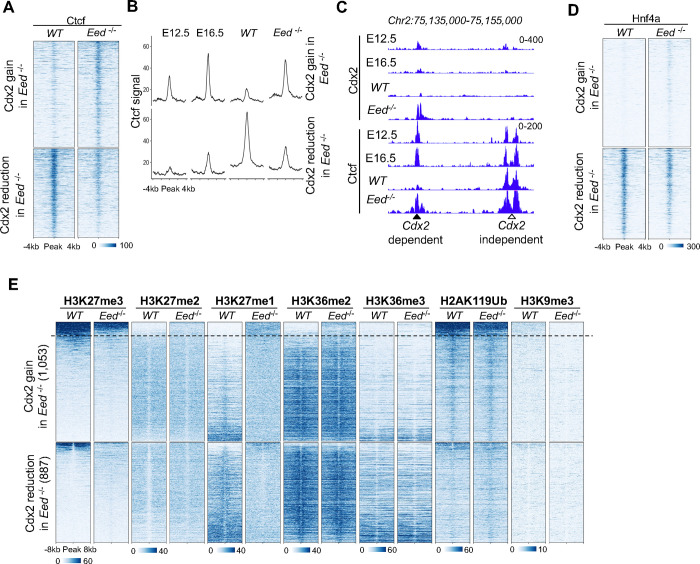
Precise spatiotemporal and cell type-specific gene expression is essential for proper tissue development and function. Transcription factors (TFs) guide this process by binding to developmental stage-specific targets and establishing an appropriate enhancer landscape. In turn, DNA and chromatin modifications direct the genomic binding of TFs. However, how TFs navigate various chromatin features and selectively bind a small portion of the millions of possible genomic target loci is still not well understood. Here we show that Cdx2 - a pioneer TF that binds distinct targets in developing versus adult intestinal epithelial cells - has a preferential affinity for a non-canonical CpG-containing motif in vivo. A higher frequency of this motif at embryonic and fetal Cdx2 target loci and the specifically methylated state of the CpG during development allows selective Cdx2 binding and activation of developmental enhancers and linked genes. Conversely, demethylation at these enhancers prohibits ectopic Cdx2 binding in adult cells, where Cdx2 binds its canonical motif without a CpG. This differential Cdx2 binding allows for corecruitment of Ctcf and Hnf4, facilitating the establishment of intestinal superenhancers during development and enhancers mediating adult homeostatic functions, respectively. Induced gain of DNA methylation in the adult mouse epithelium or cultured cells causes ectopic recruitment of Cdx2 to the developmental target loci and facilitates cobinding of the partner TFs. Together, our results demonstrate that the differential CpG motif requirements for Cdx2 binding to developmental versus adult target sites allow it to navigate different DNA methylation profiles and activate cell type-specific genes at appropriate times.
Figures




Similar articles
-
Motif distribution and DNA methylation underlie distinct Cdx2 binding during development and homeostasis.Nat Commun. 2025 Jan 22;16(1):929. doi: 10.1038/s41467-025-56187-0. Nat Commun. 2025. PMID: 39843425 Free PMC article.
-
Follistatin supplementation induces changes in CDX2 CpG methylation and improves in vitro development of bovine SCNT preimplantation embryos.Reprod Biol Endocrinol. 2021 Sep 13;19(1):141. doi: 10.1186/s12958-021-00829-7. Reprod Biol Endocrinol. 2021. PMID: 34517901 Free PMC article.
-
Transcription factors GATA4 and HNF4A control distinct aspects of intestinal homeostasis in conjunction with transcription factor CDX2.J Biol Chem. 2015 Jan 16;290(3):1850-60. doi: 10.1074/jbc.M114.620211. Epub 2014 Dec 8. J Biol Chem. 2015. PMID: 25488664 Free PMC article.
-
Activation of enhancer elements by the homeobox gene Cdx2 is cell line specific.Nucleic Acids Res. 1997 Jun 15;25(12):2293-300. doi: 10.1093/nar/25.12.2293. Nucleic Acids Res. 1997. PMID: 9171078 Free PMC article.
-
Gene methylation in gastric cancer.Clin Chim Acta. 2013 Sep 23;424:53-65. doi: 10.1016/j.cca.2013.05.002. Epub 2013 May 10. Clin Chim Acta. 2013. PMID: 23669186 Review.
References
-
- Bird A., Taggart M., Frommer M., Miller O.J., and Macleod D. (1985). A fraction of the mouse genome that is derived from islands of nonmethylated, CpG-rich DNA. Cell 40, 91–99. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous