

This is a preprint.
Metformin reduces the clonal fitness of Dnmt3a R878H hematopoietic stem and progenitor cells by reversing their aberrant metabolic and epigenetic state
- PMID: 38405837
- PMCID: PMC10889081
- DOI: 10.21203/rs.3.rs-3874821/v1
Metformin reduces the clonal fitness of Dnmt3a R878H hematopoietic stem and progenitor cells by reversing their aberrant metabolic and epigenetic state
Update in
-
Metformin reduces the competitive advantage of Dnmt3aR878H HSPCs.Nature. 2025 Jun;642(8067):421-430. doi: 10.1038/s41586-025-08871-w. Epub 2025 Apr 16. Nature. 2025. PMID: 40240595
Abstract
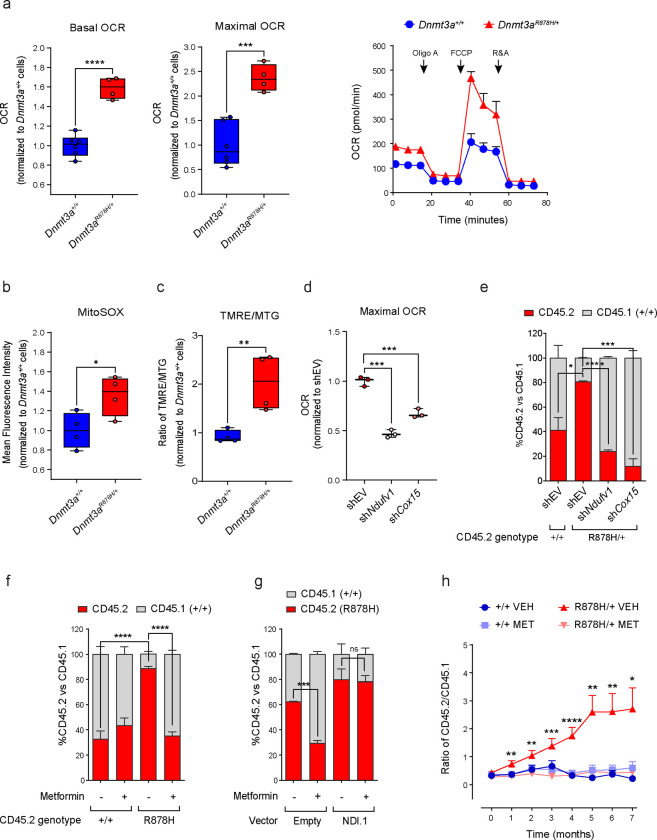
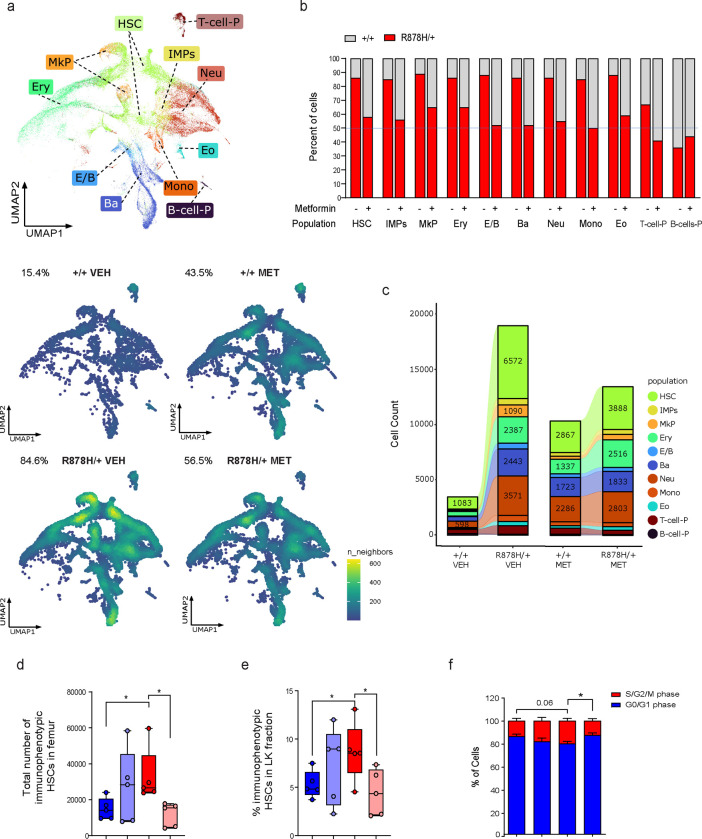
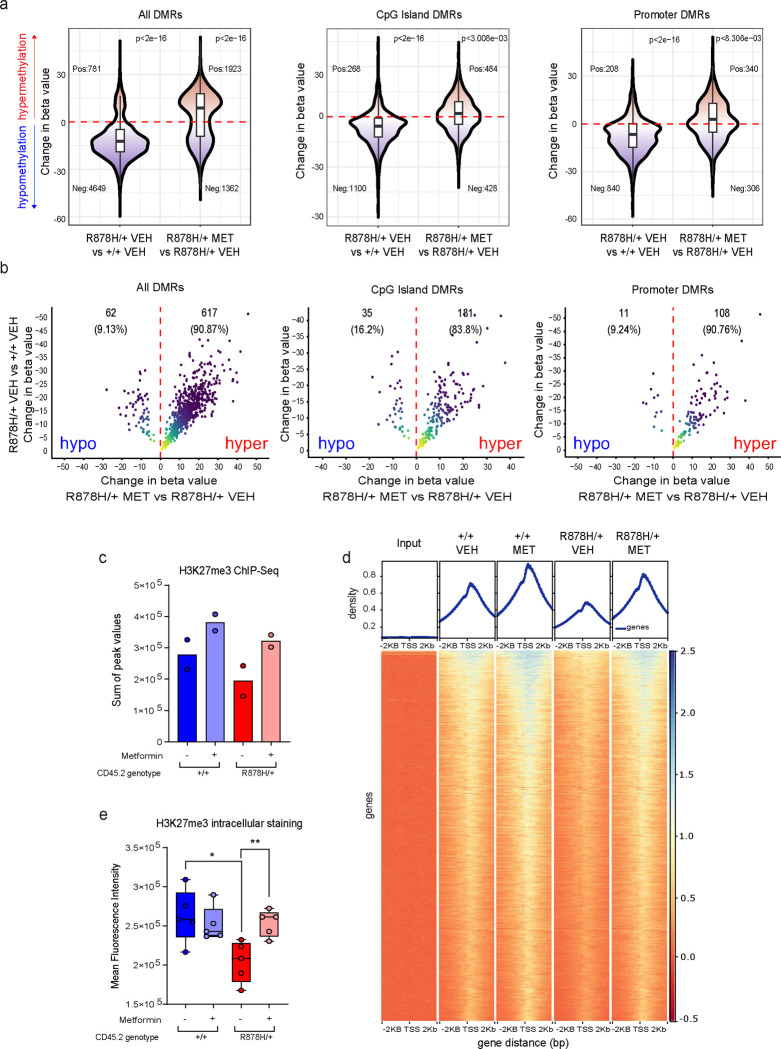
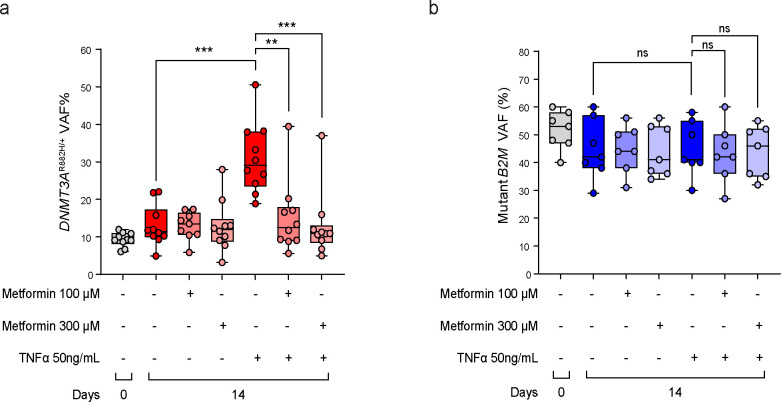
Clonal hematopoiesis (CH) arises when a hematopoietic stem cell (HSC) acquires a mutation that confers a competitive advantage over wild-type (WT) HSCs, resulting in its clonal expansion. Individuals with CH are at an increased risk of developing hematologic neoplasms and a range of age-related inflammatory illnesses1-3. Therapeutic interventions that suppress the expansion of mutant HSCs have the potential to prevent these CH-related illnesses; however, such interventions have not yet been identified. The most common CH driver mutations are in the DNA methyltransferase 3 alpha (DNMT3A) gene with arginine 882 (R882) being a mutation hotspot. Here we show that murine hematopoietic stem and progenitor cells (HSPCs) carrying the Dnmt3a R878H/+ mutation, which is equivalent to human DNMT3A R882H/+, have increased mitochondrial respiration compared with WT cells and are dependent on this metabolic reprogramming for their competitive advantage. Importantly, treatment with metformin, an oral anti-diabetic drug with inhibitory activity against complex I in the electron transport chain (ETC), reduced the fitness of Dnmt3a R878H/+ HSCs. Through a multi-omics approach, we discovered that metformin acts by enhancing the methylation potential in Dnmt3a R878H/+ HSPCs and reversing their aberrant DNA CpG methylation and histone H3K27 trimethylation (H3K27me3) profiles. Metformin also reduced the fitness of human DNMT3A R882H HSPCs generated by prime editing. Our findings provide preclinical rationale for investigating metformin as a preventive intervention against illnesses associated with DNMT3A R882 mutation-driven CH in humans.
Conflict of interest statement
Conflicts of Interest S.M.C. has received research funding from the Centre for Oncology and Immunology in Hong Kong, Celgene/BMS, AbbVie Pharmaceuticals, Agios Pharmaceuticals, and Servier Laboratories. F.G. serves as a consultant for S2 Genomics Inc. A.D.S. has received research funding from Takeda Pharmaceuticals, BMS and Medivir AB, and consulting fees/honorarium from Takeda, Novartis, Jazz, and Otsuka Pharmaceuticals. A.D.S. is named on a patent application for the use of DNT cells to treat AML. A.D.S. is a member of the Medical and Scientific Advisory Board of the Leukemia and Lymphoma Society of Canada. A.D.S. holds the Ronald N. Buick Chair in Oncology Research. J.E.D. has received research funding from Celgene/BMS, and has patents licensed to Trillium Therapeutics/Pfizer.
Figures

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
