

This is a preprint.
The interplay between electron transport chain function and iron regulatory factors influences melanin formation in Cryptococcus neoformans
- PMID: 38405941
- PMCID: PMC10888943
- DOI: 10.1101/2024.02.15.580540
The interplay between electron transport chain function and iron regulatory factors influences melanin formation in Cryptococcus neoformans
Update in
-
The interplay between electron transport chain function and iron regulatory factors influences melanin formation in Cryptococcus neoformans.mSphere. 2024 May 29;9(5):e0025024. doi: 10.1128/msphere.00250-24. Epub 2024 Apr 30. mSphere. 2024. PMID: 38687055 Free PMC article.
Abstract
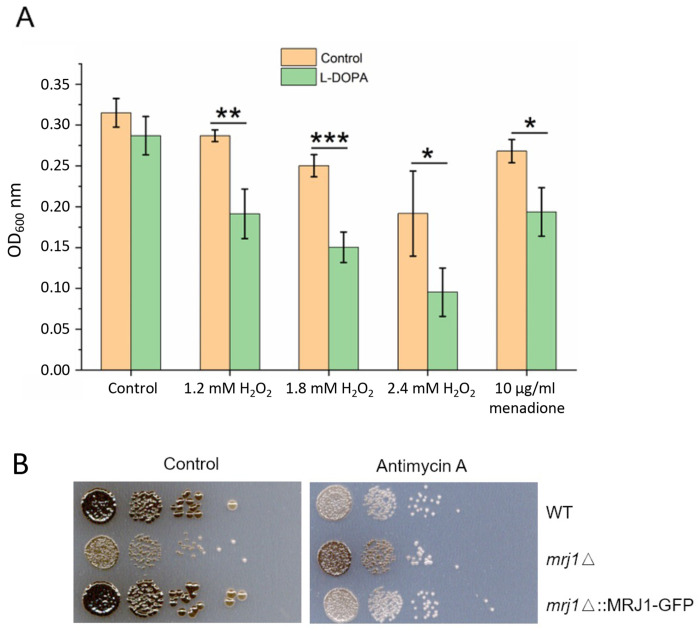
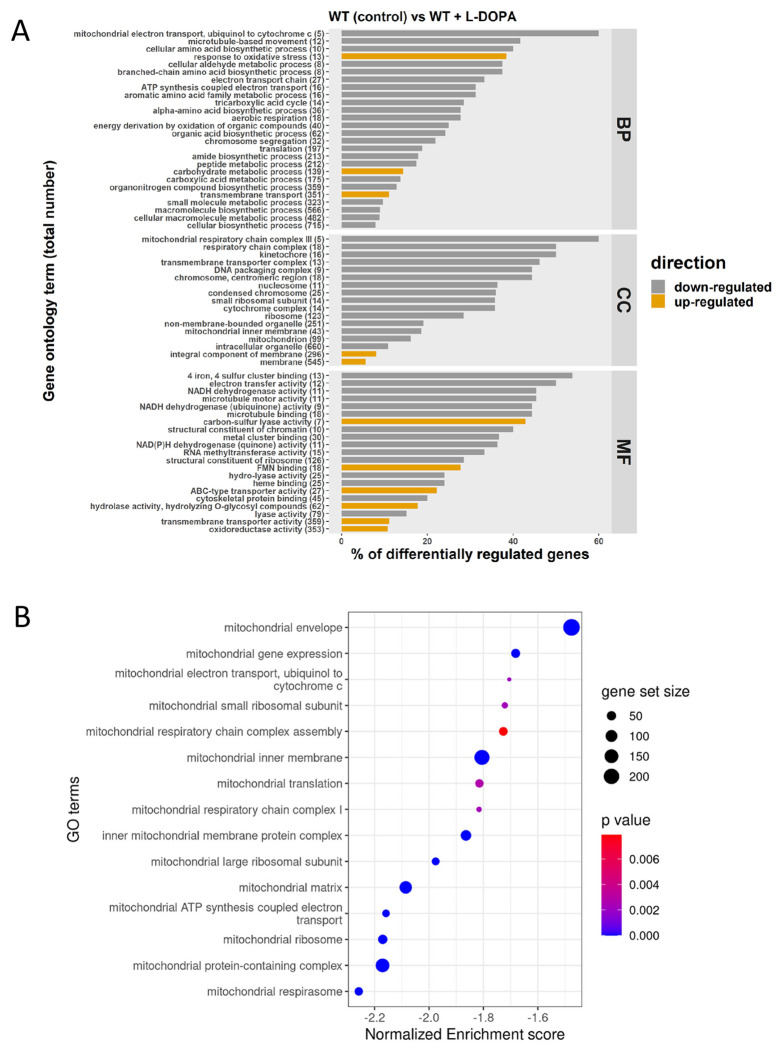
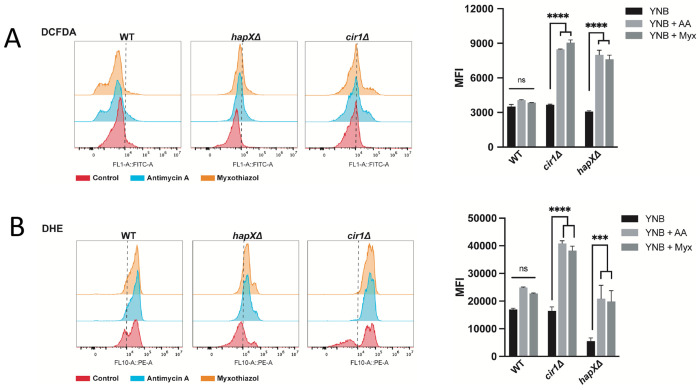
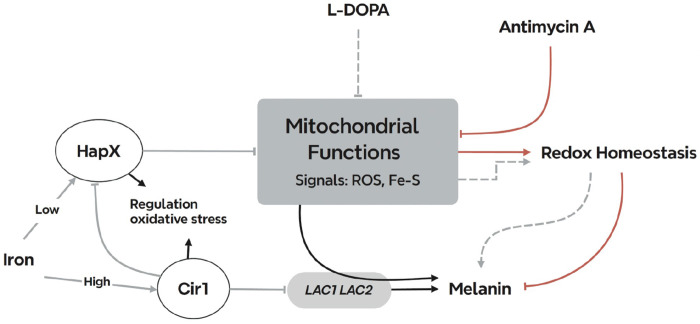
Mitochondrial functions are critical for the ability of the fungal pathogen Cryptococcus neoformans to cause disease. However, mechanistic connections between key functions such as the mitochondrial electron transport chain (ETC) and virulence factor elaboration have yet to be thoroughly characterized. Here, we observed that inhibition of ETC complex III suppressed melanin formation, a major virulence factor. This inhibition was partially blocked upon loss of Cir1 or HapX, two transcription factors that regulate iron acquisition and use. In this regard, loss of Cir1 derepresses the expression of laccase genes as a potential mechanism to restore melanin, while HapX may condition melanin formation by controlling oxidative stress. We hypothesize that ETC dysfunction alters redox homeostasis to influence melanin formation. Consistent with this idea, inhibition of growth by hydrogen peroxide was exacerbated in the presence of the melanin substrate L-DOPA. Additionally, loss of the mitochondrial chaperone Mrj1, which influences the activity of ETC complex III and reduces ROS accumulation, also partially blocked antimycin A inhibition of melanin. The phenotypic impact of mitochondrial dysfunction was consistent with RNA-Seq analyses of WT cells treated with antimycin A or L-DOPA, or cells lacking Cir1 that revealed influences on transcripts encoding mitochondrial functions (e.g., ETC components and proteins for Fe-S cluster assembly). Overall, these findings reveal mitochondria-nuclear communication via ROS and iron regulators to control virulence factor production in C. neoformans.
Keywords: RNA-Seq; electron transport chain; fungal pathogenesis; iron regulation; melanin formation; reactive oxygen species.
Conflict of interest statement
CONFLICT OF INTEREST The authors declare no competing interests.
Figures



Similar articles
-
The interplay between electron transport chain function and iron regulatory factors influences melanin formation in Cryptococcus neoformans.mSphere. 2024 May 29;9(5):e0025024. doi: 10.1128/msphere.00250-24. Epub 2024 Apr 30. mSphere. 2024. PMID: 38687055 Free PMC article.
-
A Transcriptional Regulatory Map of Iron Homeostasis Reveals a New Control Circuit for Capsule Formation in Cryptococcus neoformans.Genetics. 2020 Aug;215(4):1171-1189. doi: 10.1534/genetics.120.303270. Epub 2020 Jun 24. Genetics. 2020. PMID: 32580959 Free PMC article.
-
The Monothiol Glutaredoxin Grx4 Regulates Iron Homeostasis and Virulence in Cryptococcus neoformans.mBio. 2018 Dec 4;9(6):e02377-18. doi: 10.1128/mBio.02377-18. mBio. 2018. PMID: 30514787 Free PMC article.
-
Coordinated regulation of iron metabolism in Cryptococcus neoformans by GATA and CCAAT transcription factors: connections with virulence.Curr Genet. 2021 Aug;67(4):583-593. doi: 10.1007/s00294-021-01172-5. Epub 2021 Mar 24. Curr Genet. 2021. PMID: 33760942 Free PMC article. Review.
-
An encapsulation of iron homeostasis and virulence in Cryptococcus neoformans.Trends Microbiol. 2013 Sep;21(9):457-65. doi: 10.1016/j.tim.2013.05.007. Epub 2013 Jun 25. Trends Microbiol. 2013. PMID: 23810126 Free PMC article. Review.
References
-
- Kretschmer M, Damoo D, Sun S, Lee CW, Croll D, Brumer H, Kronstad J. 2022. Organic acids and glucose prime late-stage fungal biotrophy in maize. Science 376:1187–1191. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous