

Altered IL-7 signaling in CD4+ T cells from patients with visceral leishmaniasis
- PMID: 38408097
- PMCID: PMC10919868
- DOI: 10.1371/journal.pntd.0011960
Altered IL-7 signaling in CD4+ T cells from patients with visceral leishmaniasis
Abstract
Background: CD4+ T cells play a central role in control of L. donovani infection, through IFN-γ production required for activation of macrophages and killing of intracellular parasites. Impaired control of parasites can in part be explained by hampered CD4+ T cells effector functions in visceral leishmaniasis (VL) patients. In a recent studies that defined transcriptional signatures for CD4+ T cells from active VL patients, we found that expression of the IL-7 receptor alpha chain (IL-7Rα; CD127) was downregulated, compared to CD4+ T cells from endemic controls (ECs). Since IL-7 signaling is critical for the survival and homeostatic maintenance of CD4+ T cells, we investigated this signaling pathway in VL patients, relative to ECs.
Methods: CD4+ T cells were enriched from peripheral blood collected from VL patients and EC subjects and expression of IL7 and IL7RA mRNA was measured by real time qPCR. IL-7 signaling potential and surface expression of CD127 and CD132 on CD4+ T cell was analyzed by multicolor flow cytometry. Plasma levels of soluble IL-7 and sIL-7Rα were measured by ELISA.
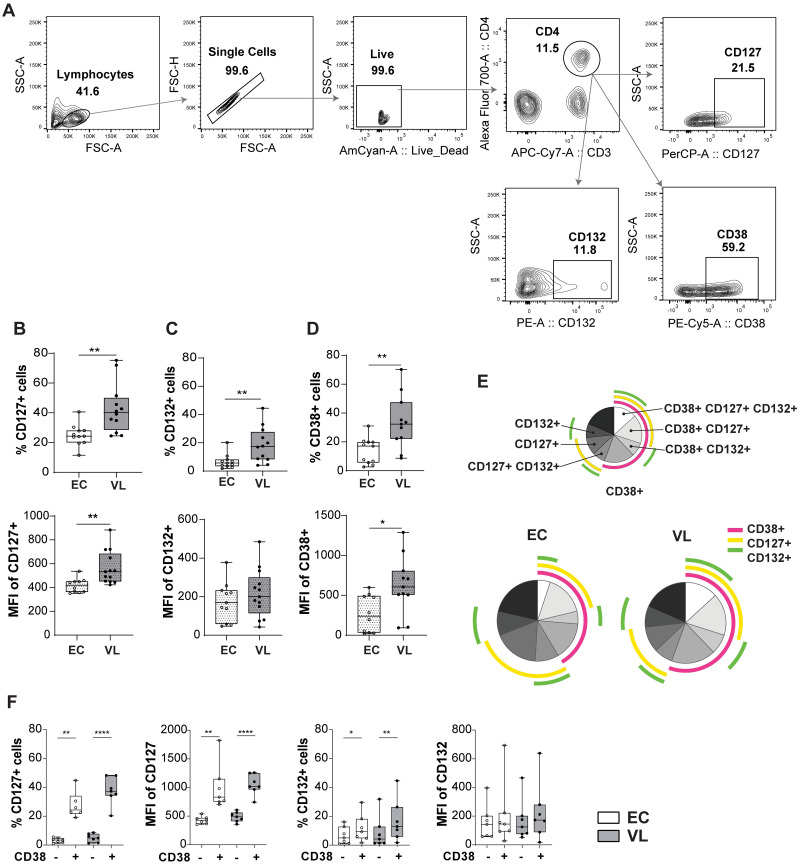
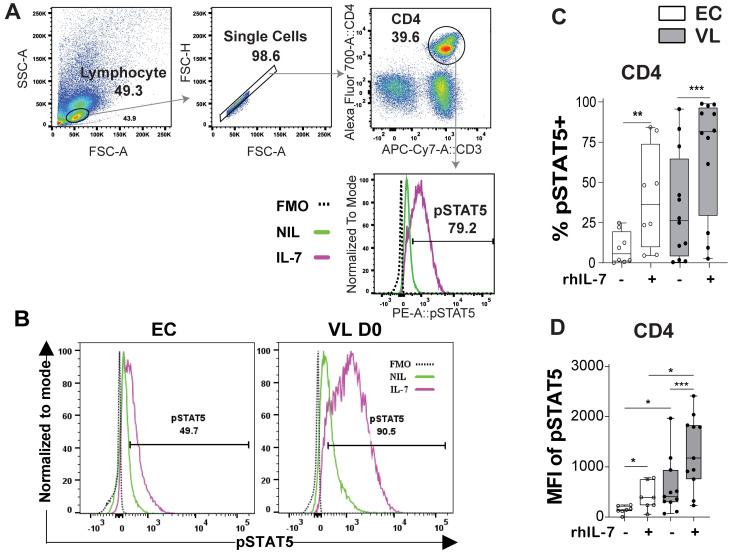
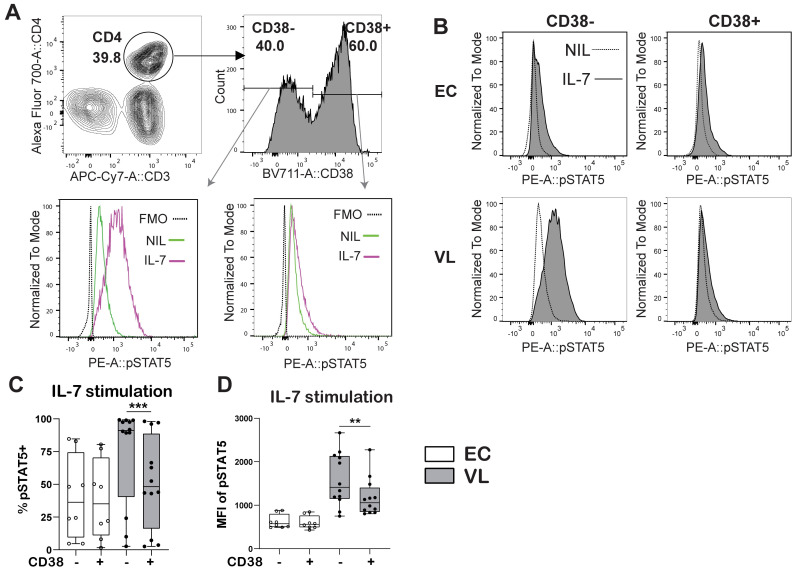
Result: Transcriptional profiling data sets generated previously from our group showed lower IL7RA mRNA expression in VL CD4+ T cells as compared to EC. A significant reduction was, however not seen when assessing IL7RA mRNA by RT-qPCR. Yet, the levels of soluble IL-7Rα (sIL-7Rα) were reduced in plasma of VL patients compared to ECs. Furthermore, the levels of soluble IL-7 were higher in plasma from VL patients compared to ECs. Interestingly, expression of the IL-7Rα protein was higher on VL patient CD4+ T cells as compared to EC, with activated CD38+ CD4+ T cells showing higher surface expression of IL-7Rα compared to CD38- CD4+ T cells in VL patients. CD4+ T cells from VL patients had higher signaling potential baseline and after stimulation with recombinant human IL-7 (rhIL-7) compared to EC, as measured by phosphorylation of STAT5 (pSTAT5). Interestingly, it was the CD38 negative cells that had the highest level of pSTAT5 in VL patient CD4+ T cells after IL-7 stimulation. Thus, despite unaltered or potentially lowered IL7RA mRNA expression by CD4+ T cells from VL patients, the surface expression of the IL-7Rα was higher compared to EC and increased pSTAT5 was seen following exposure to rhIL-7. Accordingly, IL-7 signaling appears to be functional and even enhanced in VL CD4+ T cells and cannot explain the impaired effector function of VL CD4+ T cells. The enhanced plasma IL-7 may serve as part of homeostatic feedback mechanism regulating IL7RA expression in CD4+ T cells.
Copyright: © 2024 Kumar et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



Similar articles
-
Increased bone marrow interleukin-7 (IL-7)/IL-7R levels but reduced IL-7 responsiveness in HIV-positive patients lacking CD4+ gain on antiviral therapy.PLoS One. 2010 Dec 31;5(12):e15663. doi: 10.1371/journal.pone.0015663. PLoS One. 2010. PMID: 21209878 Free PMC article.
-
Splenic accumulation of IL-10 mRNA in T cells distinct from CD4+CD25+ (Foxp3) regulatory T cells in human visceral leishmaniasis.J Exp Med. 2007 Apr 16;204(4):805-17. doi: 10.1084/jem.20061141. Epub 2007 Mar 26. J Exp Med. 2007. PMID: 17389235 Free PMC article.
-
Multifunctional, TNF-α and IFN-γ-Secreting CD4 and CD8 T Cells and CD8High T Cells Are Associated With the Cure of Human Visceral Leishmaniasis.Front Immunol. 2021 Oct 28;12:773983. doi: 10.3389/fimmu.2021.773983. eCollection 2021. Front Immunol. 2021. PMID: 34777391 Free PMC article.
-
Reduced numbers of IL-7 receptor (CD127) expressing immune cells and IL-7-signaling defects in peripheral blood from patients with breast cancer.Int J Cancer. 2007 Oct 1;121(7):1512-9. doi: 10.1002/ijc.22854. Int J Cancer. 2007. PMID: 17546596
-
IL7RA genetic variants differentially affect IL-7Rα expression and alternative splicing: a role in autoimmune and infectious diseases?Genes Immun. 2020 Feb;21(2):83-90. doi: 10.1038/s41435-019-0091-y. Epub 2020 Jan 13. Genes Immun. 2020. PMID: 31929513 Review.
Cited by
-
Chemokines Signature and T Cell Dynamics in Leishmaniasis: Molecular insight and therapeutic application.Expert Rev Mol Med. 2024 Nov 26;27:1-55. doi: 10.1017/erm.2024.36. Online ahead of print. Expert Rev Mol Med. 2024. PMID: 39587036 Free PMC article. Review.
References
-
- Leishmaniasis. [cited 25 Jul 2023]. https://www.who.int/news-room/fact-sheets/detail/leishmaniasis.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
