

Precise fine-turning of GhTFL1 by base editing tools defines ideal cotton plant architecture
- PMID: 38409014
- PMCID: PMC10895741
- DOI: 10.1186/s13059-024-03189-8
Precise fine-turning of GhTFL1 by base editing tools defines ideal cotton plant architecture
Abstract
Background: CRISPR/Cas-derived base editor enables precise editing of target sites and has been widely used for basic research and crop genetic improvement. However, the editing efficiency of base editors at different targets varies greatly.
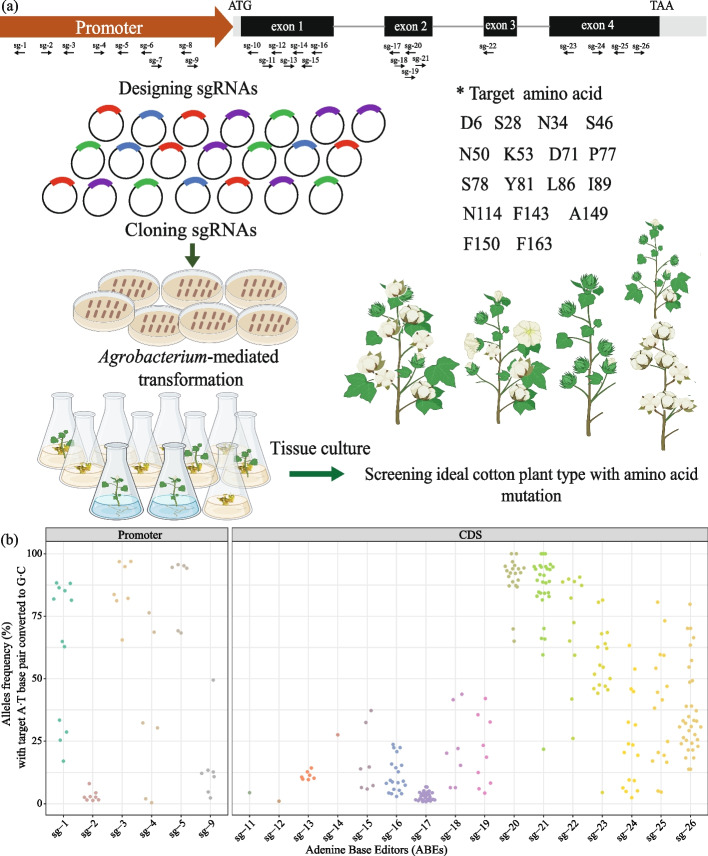
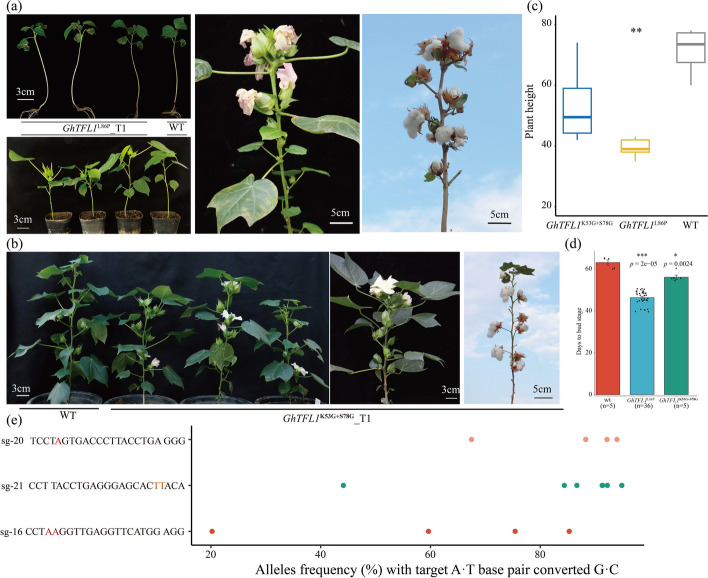
Results: Here, we develop a set of highly efficient base editors in cotton plants. GhABE8e, which is fused to conventional nCas9, exhibits 99.9% editing efficiency, compared to GhABE7.10 with 64.9%, and no off-target editing is detected. We further replace nCas9 with dCpf1, which recognizes TTTV PAM sequences, to broaden the range of the target site. To explore the functional divergence of TERMINAL FLOWER 1 (TFL1), we edit the non-coding and coding regions of GhTFL1 with 26 targets to generate a comprehensive allelic population including 300 independent lines in cotton. This allows hidden pleiotropic roles for GhTFL1 to be revealed and allows us to rapidly achieve directed domestication of cotton and create ideotype germplasm with moderate height, shortened fruiting branches, compact plant, and early-flowering. Further, by exploring the molecular mechanism of the GhTFL1L86P and GhTFL1K53G+S78G mutations, we find that the GhTFL1L86P mutation weakens the binding strength of the GhTFL1 to other proteins but does not lead to a complete loss of GhTFL1 function.
Conclusions: This strategy provides an important technical platform and genetic information for the study and creation of ideal plant architecture.
Keywords: Base editor; Cotton; Directed evolution; Genome editing; Plant ideotype.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures




References
-
- Yu H, Lin T, Meng X, Du H, Zhang J, Liu G, Chen M, Jing Y, Kou L, Li X, et al. A route to de novo domestication of wild allotetraploid rice. Cell. 2021;184:1156–1170.e1114. - PubMed
-
- Soyk S, Müller NA, Park SJ, Schmalenbach I, Jiang K, Hayama R, Zhang L, Van Eck J, Jiménez-Gómez JM, Lippman ZB. Variation in the flowering gene SELF PRUNING 5G promotes day-neutrality and early yield in tomato. Nat Genet. 2017;49:162–168. - PubMed
-
- Wang JY, Doudna JA. CRISPR technology: a decade of genome editing is only the beginning. Science. 2023;379:eadd8643. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
