

Human cytomegalovirus exploits STING signaling and counteracts IFN/ISG induction to facilitate infection of dendritic cells
- PMID: 38409141
- PMCID: PMC10897438
- DOI: 10.1038/s41467-024-45614-3
Human cytomegalovirus exploits STING signaling and counteracts IFN/ISG induction to facilitate infection of dendritic cells
Abstract
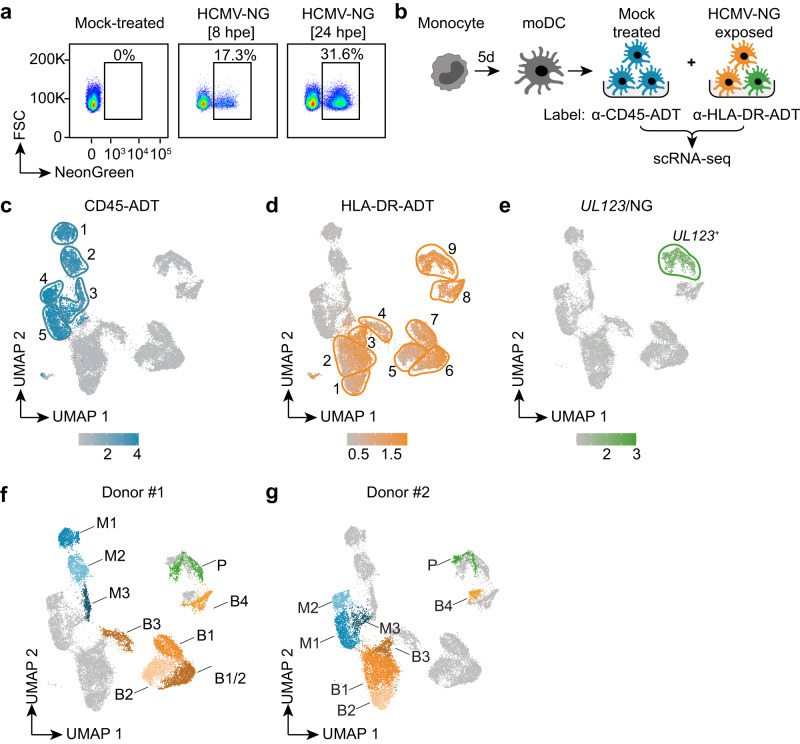
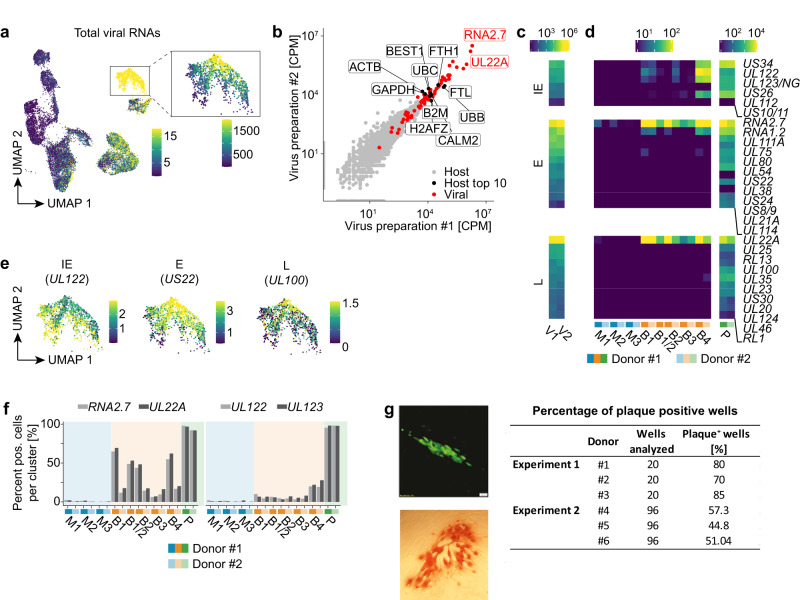
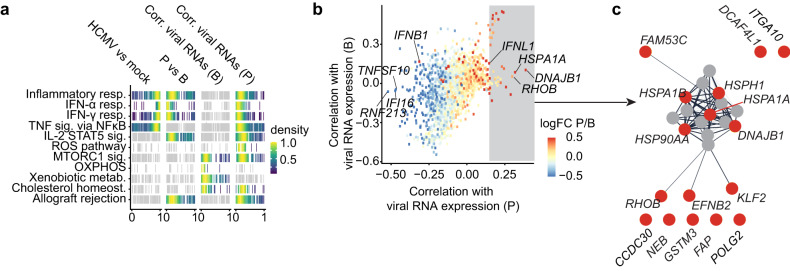
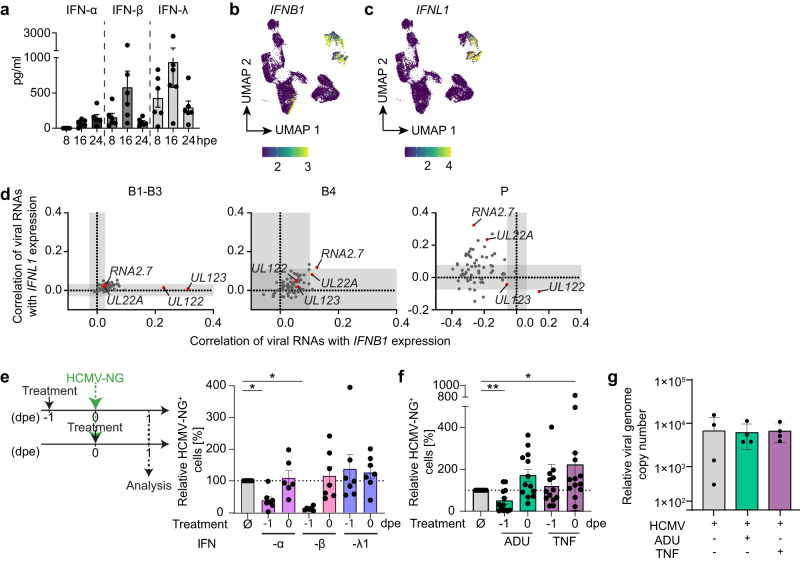
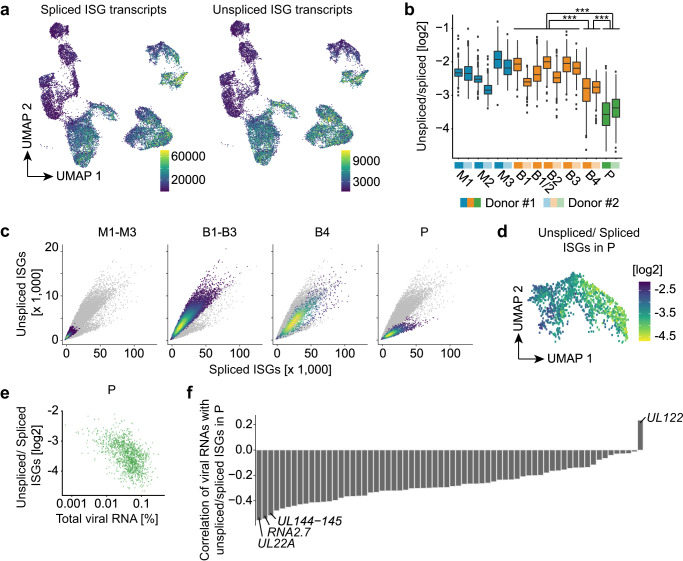
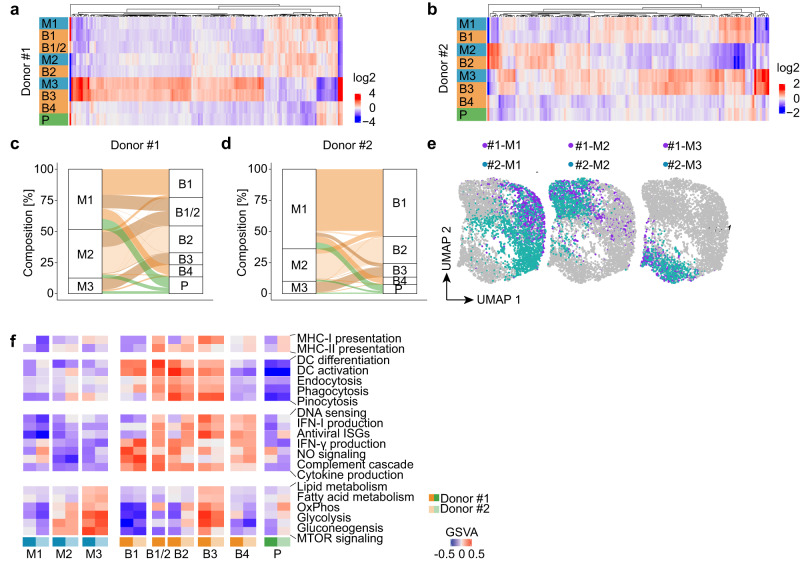
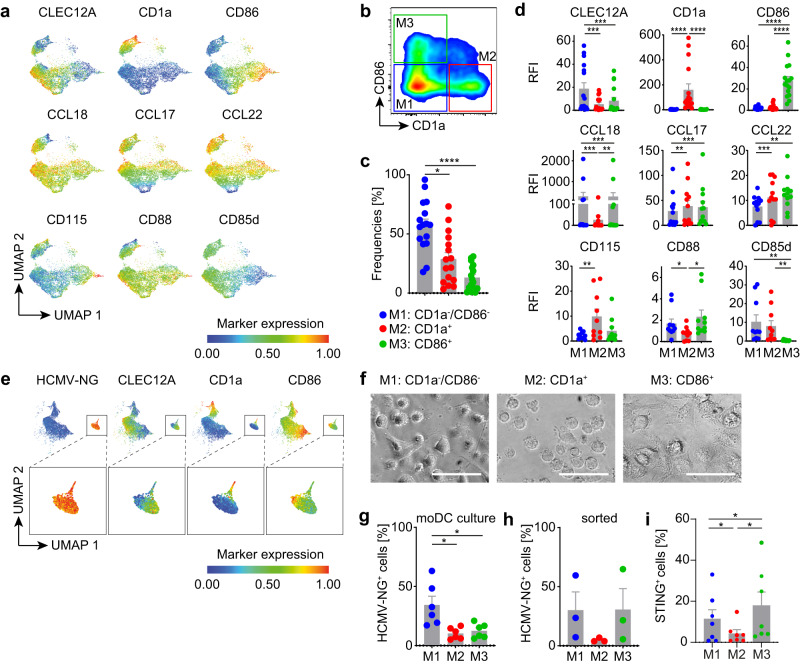
Human cytomegalovirus (HCMV) is a widespread pathogen that in immunocompromised hosts can cause life-threatening disease. Studying HCMV-exposed monocyte-derived dendritic cells by single-cell RNA sequencing, we observe that most cells are entered by the virus, whereas less than 30% of them initiate viral gene expression. Increased viral gene expression is associated with activation of the stimulator of interferon genes (STING) that usually induces anti-viral interferon responses, and with the induction of several pro- (RHOB, HSP1A1, DNAJB1) and anti-viral (RNF213, TNFSF10, IFI16) genes. Upon progression of infection, interferon-beta but not interferon-lambda transcription is inhibited. Similarly, interferon-stimulated gene expression is initially induced and then shut off, thus further promoting productive infection. Monocyte-derived dendritic cells are composed of 3 subsets, with one being especially susceptible to HCMV. In conclusion, HCMV permissiveness of monocyte-derived dendritic cells depends on complex interactions between virus sensing, regulation of the interferon response, and viral gene expression.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- WHO, CDC & International Clearinghouse for Birth Defects. Birth defects surveillance: a manual for programme managers. 2nd edition https://www.who.int/publications/i/item/9789240015395 (2020).
-
- Taylor-Wiedeman J, Sissons JG, Borysiewicz LK, Sinclair JH. Monocytes are a major site of persistence of human cytomegalovirus in peripheral blood mononuclear cells. J. Gen. Virol. 1991;72:2059–2064. - PubMed
-
- Sinzger C, Plachter B, Grefte A, The TH, Jahn G. Tissue macrophages are infected by human cytomegalovirus in vivo. J. Infect. Dis. 1996;173:240–245. - PubMed
-
- Spector WG, Walters MN, Willoughby DA. The origin of the mononuclear cells in inflammatory exudates induced by fibrinogen. J. Pathol. Bacteriol. 1965;90:181–192. - PubMed
MeSH terms
Substances
Grants and funding
- 158989968 - SFB 900-B2/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 398367752 - FOR 2830/Deutsche Forschungsgemeinschaft (German Research Foundation)
- EXC 2155 "RESIST" - Project ID 39087428/Deutsche Forschungsgemeinschaft (German Research Foundation)
- DO 1275/7-1/Deutsche Forschungsgemeinschaft (German Research Foundation)
- ER 927/2-1 - FOR2830/Deutsche Forschungsgemeinschaft (German Research Foundation)
- COALITION/Niedersächsisches Ministerium für Wissenschaft und Kultur (Ministry for Science and Culture of Lower Saxony)
- Marie Skłodowska-Curie Actions Innovative Training Network (VIROINF: 955974)/European Commission (EC)
- Marie Skłodowska-Curie Actions Innovative Training Network (VIROINF: 955974)/European Commission (EC)
- 0703/68674/5/2017/Bayerisches Staatsministerium für Wirtschaft und Medien, Energie und Technologie (Bavarian Ministry of Economic Affairs and Media, Energy and Technology)
- 0703/89374/3/2017/Bayerisches Staatsministerium für Wirtschaft und Medien, Energie und Technologie (Bavarian Ministry of Economic Affairs and Media, Energy and Technology)
- 0703/68674/5/2017/Bayerisches Staatsministerium für Wirtschaft und Medien, Energie und Technologie (Bavarian Ministry of Economic Affairs and Media, Energy and Technology)
- 0703/89374/3/2017/Bayerisches Staatsministerium für Wirtschaft und Medien, Energie und Technologie (Bavarian Ministry of Economic Affairs and Media, Energy and Technology)
- 0703/68674/5/2017/Bayerisches Staatsministerium für Wirtschaft und Medien, Energie und Technologie (Bavarian Ministry of Economic Affairs and Media, Energy and Technology)
- 0703/89374/3/2017/Bayerisches Staatsministerium für Wirtschaft und Medien, Energie und Technologie (Bavarian Ministry of Economic Affairs and Media, Energy and Technology)
LinkOut - more resources
Full Text Sources
Research Materials
