

Cerebellar interpositus nucleus exhibits time-dependent errors and predictive responses
- PMID: 38409163
- PMCID: PMC10897197
- DOI: 10.1038/s41539-024-00224-y
Cerebellar interpositus nucleus exhibits time-dependent errors and predictive responses
Abstract
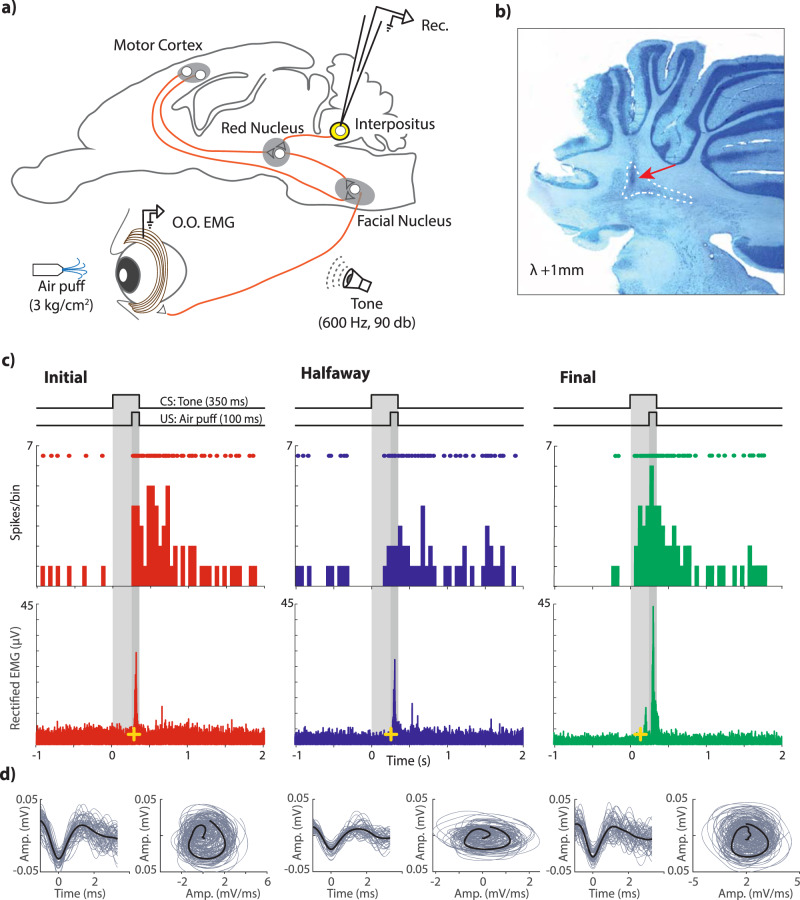
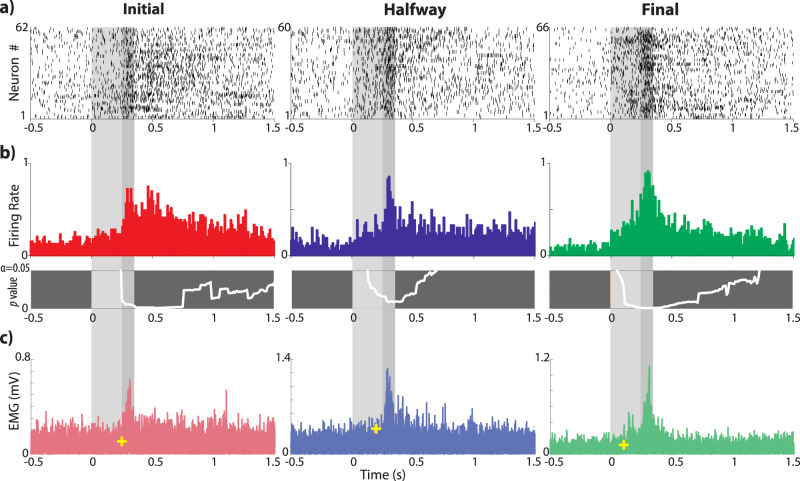
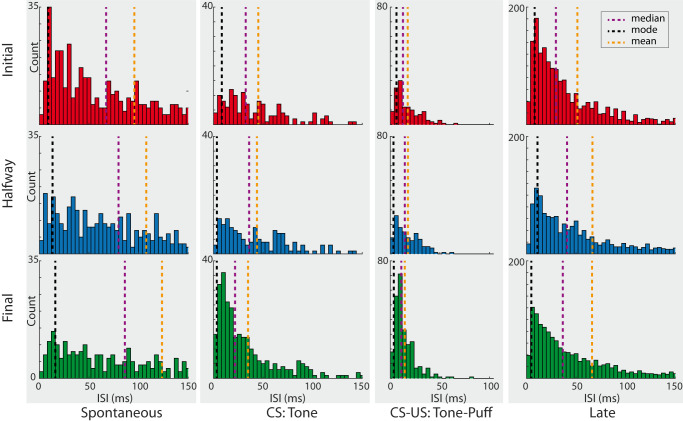
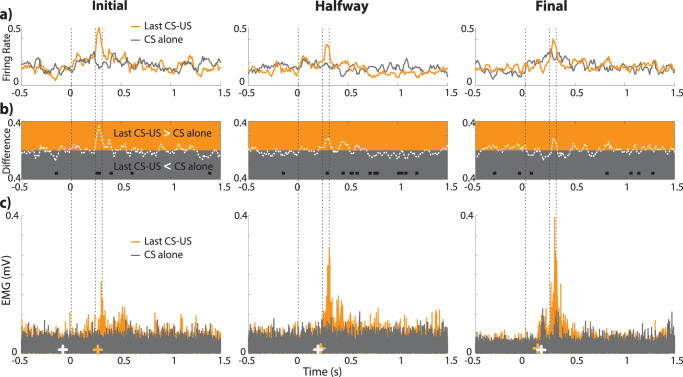
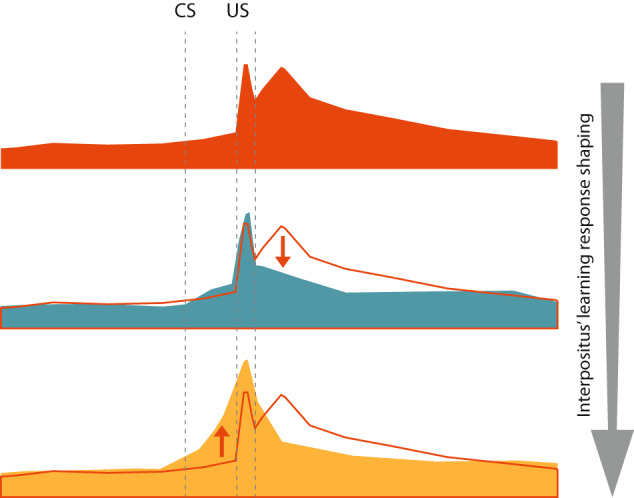
Learning is a functional state of the brain that should be understood as a continuous process, rather than being restricted to the very moment of its acquisition, storage, or retrieval. The cerebellum operates by comparing predicted states with actual states, learning from errors, and updating its internal representation to minimize errors. In this regard, we studied cerebellar interpositus nucleus (IPn) functional capabilities by recording its unitary activity in behaving rabbits during an associative learning task: the classical conditioning of eyelid responses. We recorded IPn neurons in rabbits during classical eyeblink conditioning using a delay paradigm. We found that IPn neurons reduce error signals across conditioning sessions, simultaneously increasing and transmitting spikes before the onset of the unconditioned stimulus. Thus, IPn neurons generate predictions that optimize in time and shape the conditioned eyeblink response. Our results are consistent with the idea that the cerebellum works under Bayesian rules updating the weights using the previous history.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
The Motor Cortex Is Involved in the Generation of Classically Conditioned Eyelid Responses in Behaving Rabbits.J Neurosci. 2016 Jun 29;36(26):6988-7001. doi: 10.1523/JNEUROSCI.4190-15.2016. J Neurosci. 2016. PMID: 27358456 Free PMC article.
-
The Anatomy and Physiology of Eyeblink Classical Conditioning.Curr Top Behav Neurosci. 2018;37:297-323. doi: 10.1007/7854_2016_455. Curr Top Behav Neurosci. 2018. PMID: 28025812 Review.
-
Neuronal responses of the rabbit cerebellum during acquisition and performance of a classically conditioned nictitating membrane-eyelid response.J Neurosci. 1984 Nov;4(11):2811-22. doi: 10.1523/JNEUROSCI.04-11-02811.1984. J Neurosci. 1984. PMID: 6502205 Free PMC article.
-
Dissociaton of conditioned eye and limb responses in the cerebellar interpositus.Physiol Behav. 2007 May 16;91(1):9-14. doi: 10.1016/j.physbeh.2007.01.006. Epub 2007 Jan 20. Physiol Behav. 2007. PMID: 17320121
-
The role of interpositus nucleus in eyelid conditioned responses.Cerebellum. 2002 Dec;1(4):289-308. doi: 10.1080/147342202320883597. Cerebellum. 2002. PMID: 12879967 Review.
Cited by
-
Direct brain recordings reveal implicit encoding of structure in random auditory streams.Sci Rep. 2025 Apr 27;15(1):14725. doi: 10.1038/s41598-025-98865-5. Sci Rep. 2025. PMID: 40289162 Free PMC article.
References
-
- Delgado-García JM, Gruart A. The role of interpositus nucleus in eyelid conditioned responses. Cerebellum. 2002;1:289–308. - PubMed
-
- Delgado-García JM, Gruart A. Learning as a functional state of the brain: Studies in wild-type and transgenic animals. Adv. Exp. Med. Biol. 2017;1015:75–93. - PubMed
-
- Clark A. Whatever next? Predictive brains, situated agents, and the future of cognitive science. Behav. Brain Sci. 2013;36:181–204. - PubMed
Grants and funding
- DOC-00309/Consejería de Economía, Innovación, Ciencia y Empleo, Junta de Andalucía (Ministry of Economy, Innovation, Science and Employment, Government of Andalucia)
- BIO-122/Consejería de Economía, Innovación, Ciencia y Empleo, Junta de Andalucía (Ministry of Economy, Innovation, Science and Employment, Government of Andalucia)
- PID2021-122446NB-100/Ministerio de Economía y Competitividad (Ministry of Economy and Competitiveness)
LinkOut - more resources
Full Text Sources
