

Siderophores promote cooperative interspecies and intraspecies cross-protection against antibiotics in vitro
- PMID: 38409256
- PMCID: PMC11239084
- DOI: 10.1038/s41564-024-01601-4
Siderophores promote cooperative interspecies and intraspecies cross-protection against antibiotics in vitro
Abstract
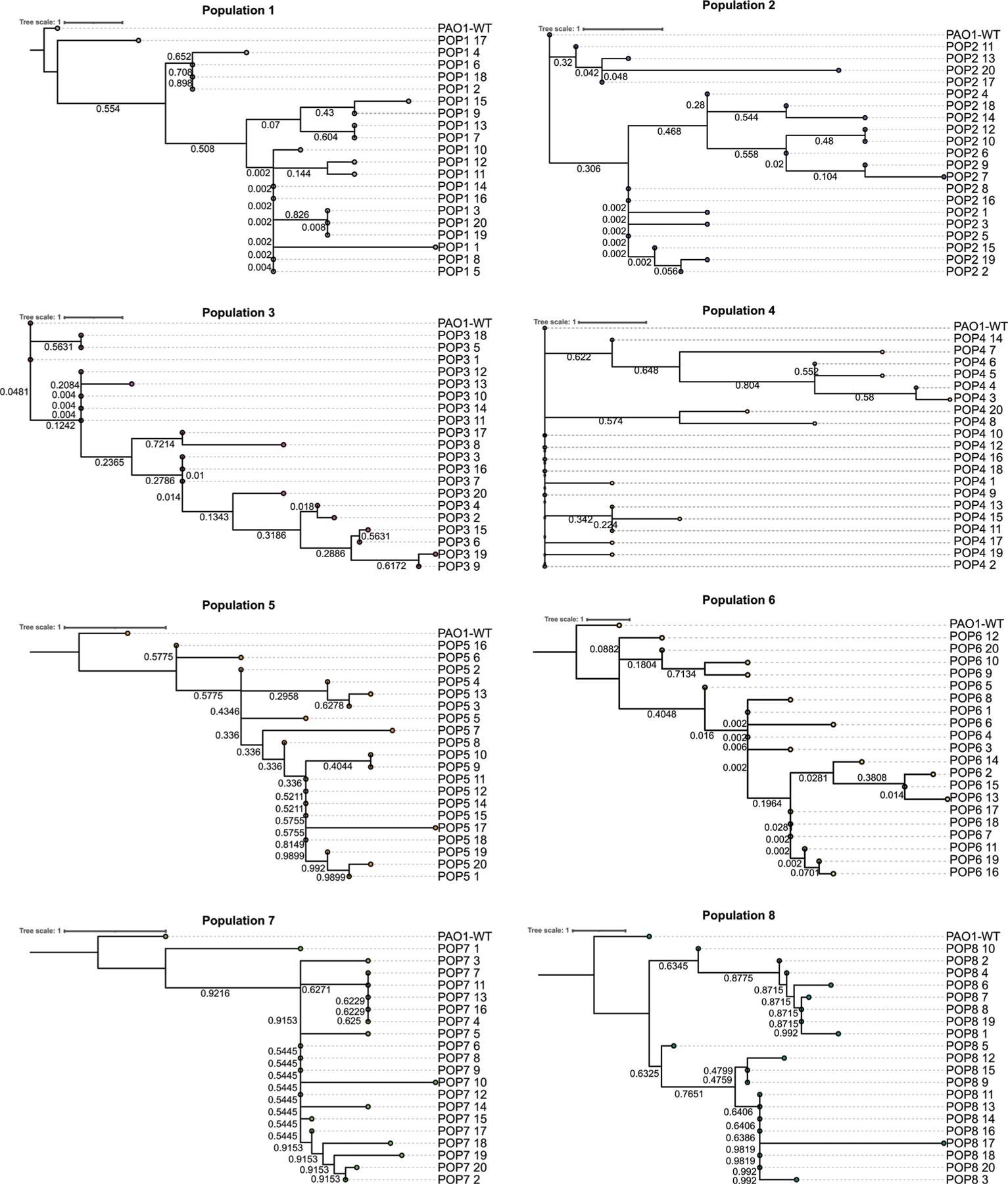
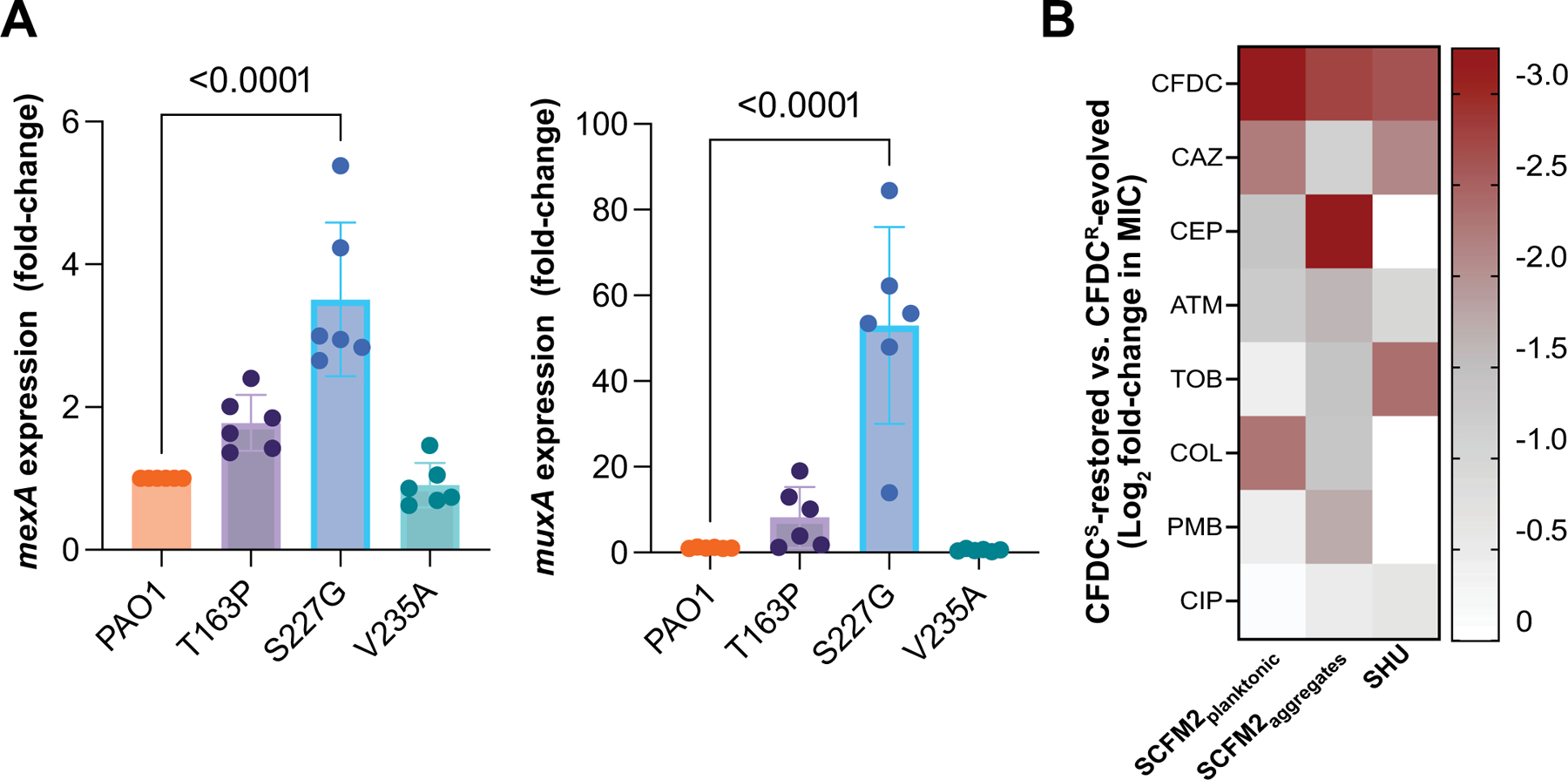
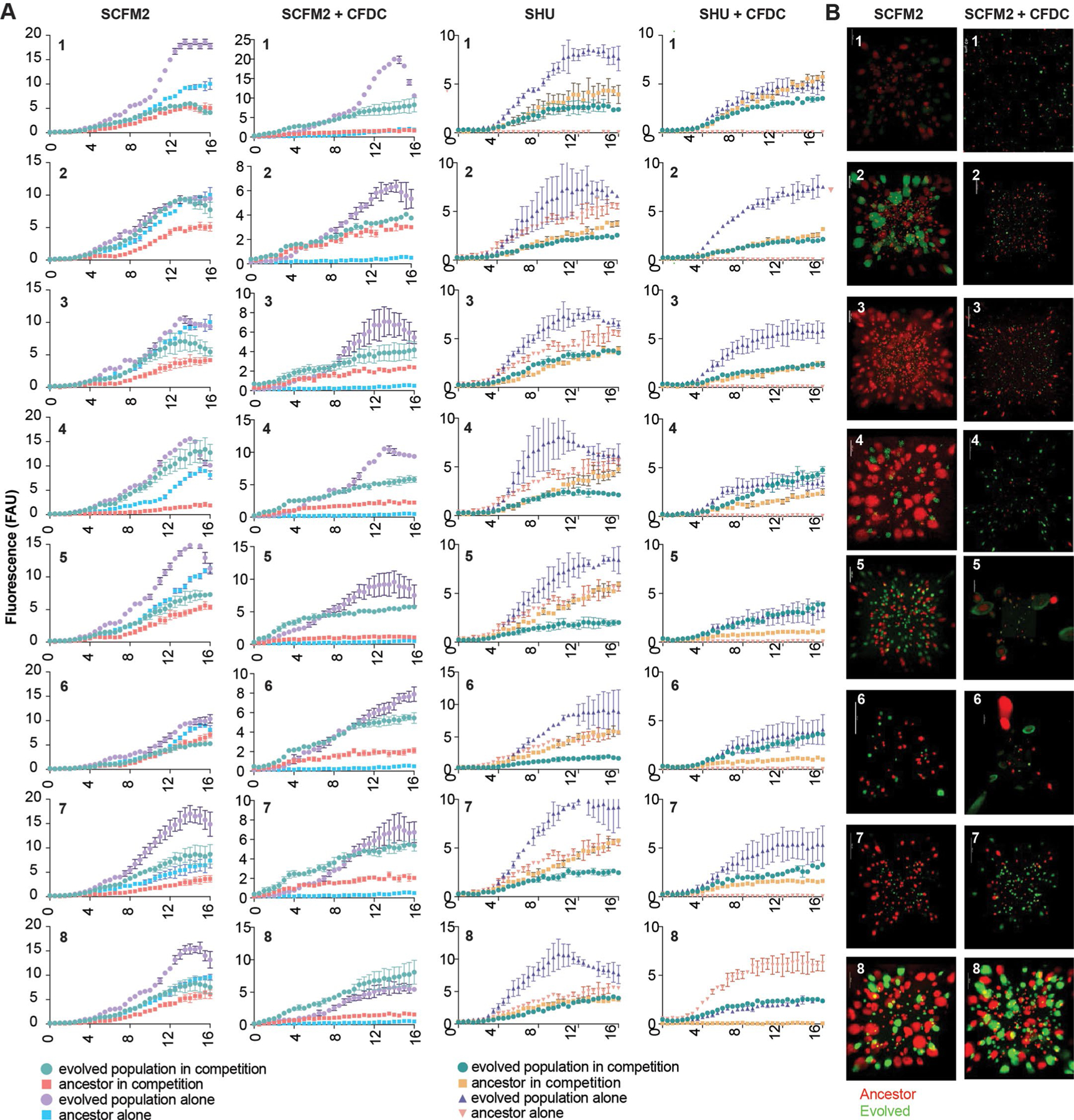
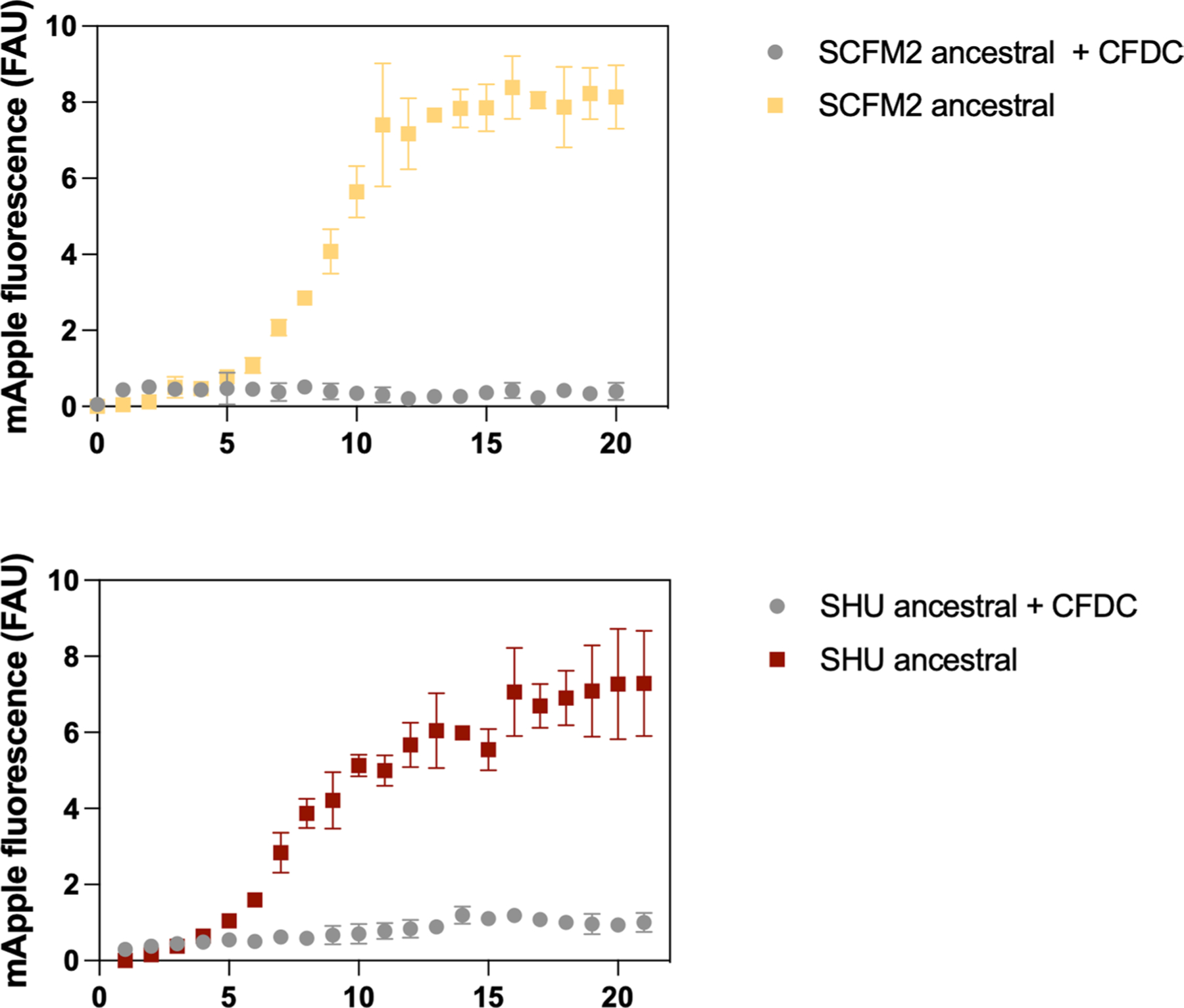
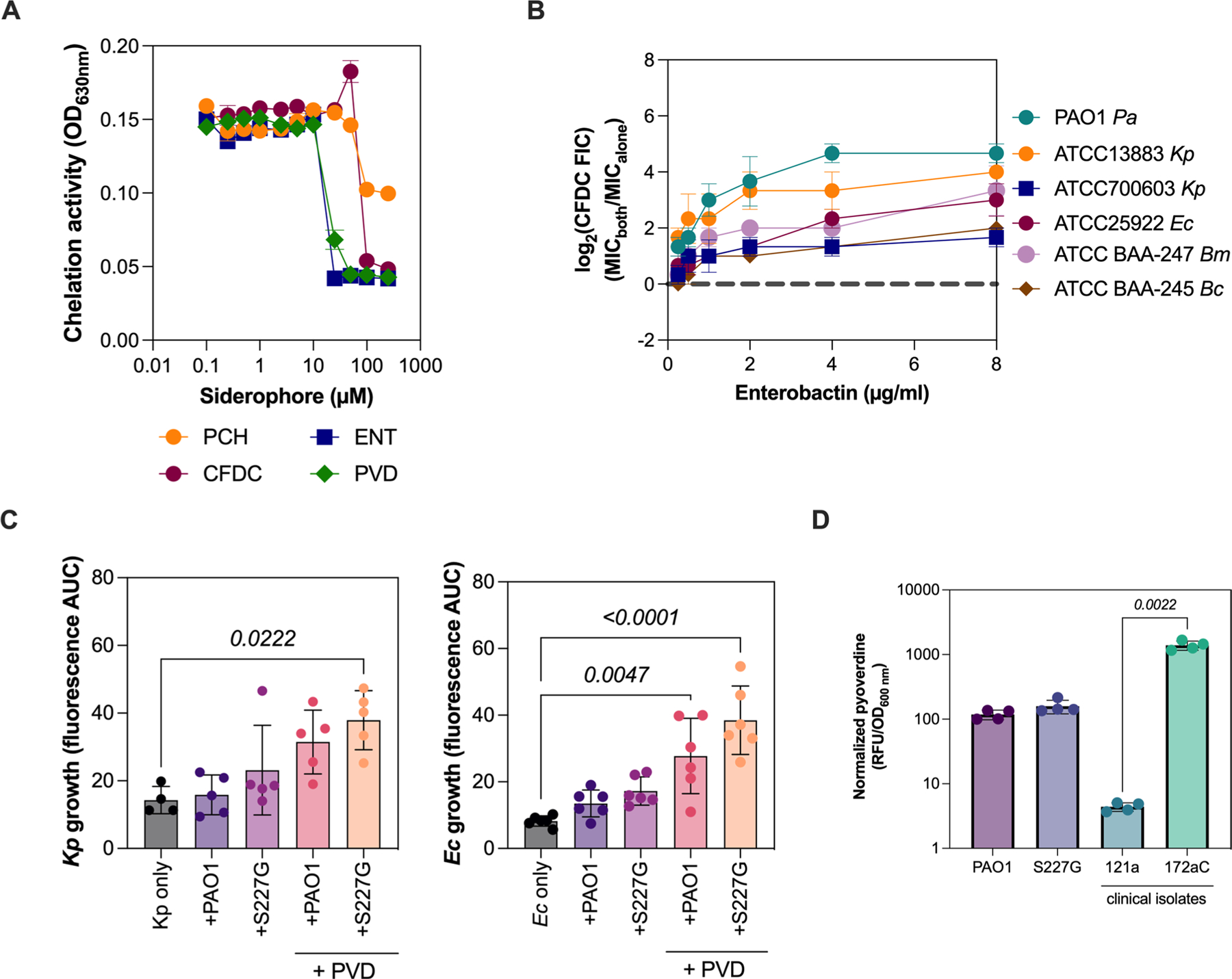
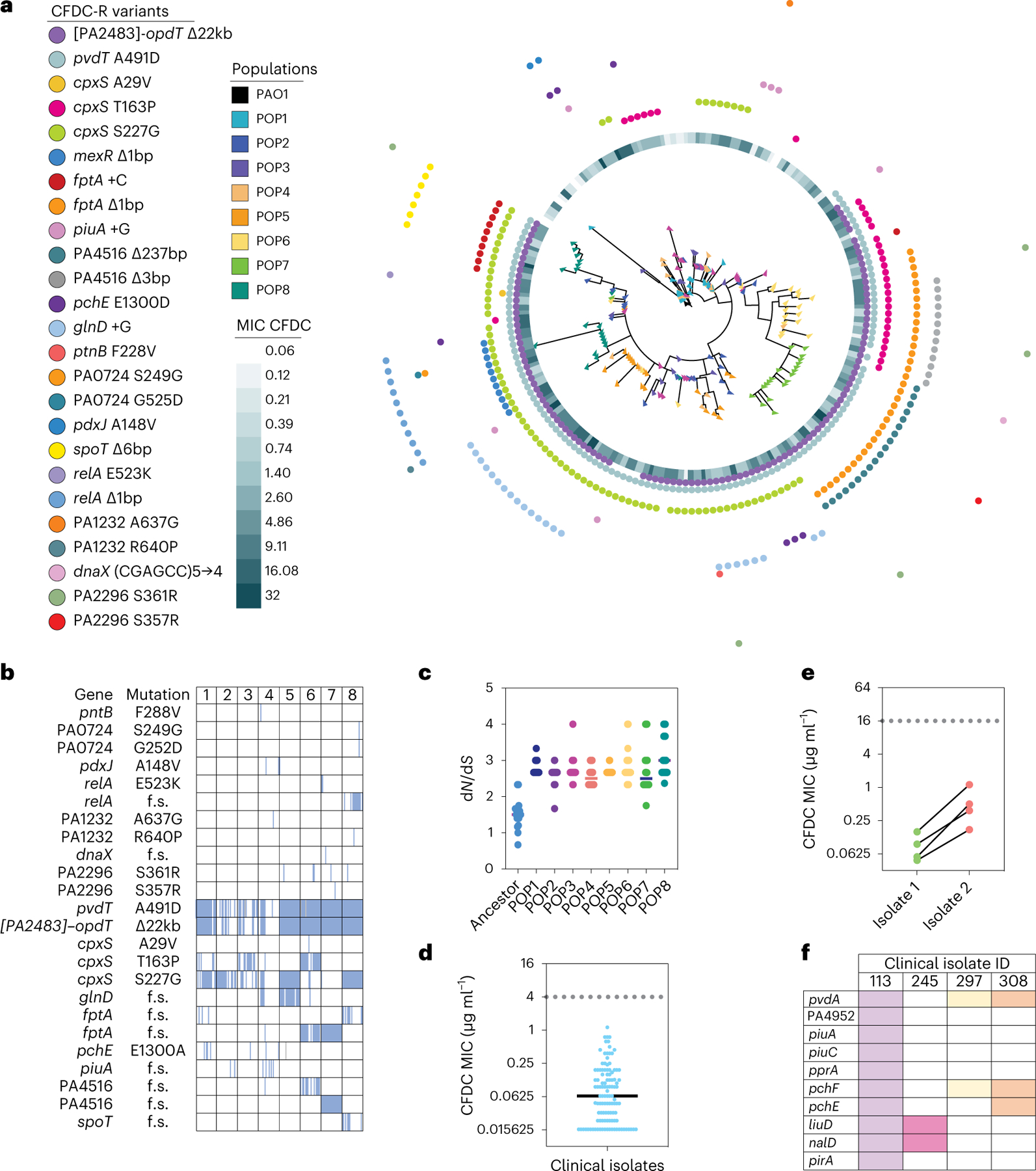
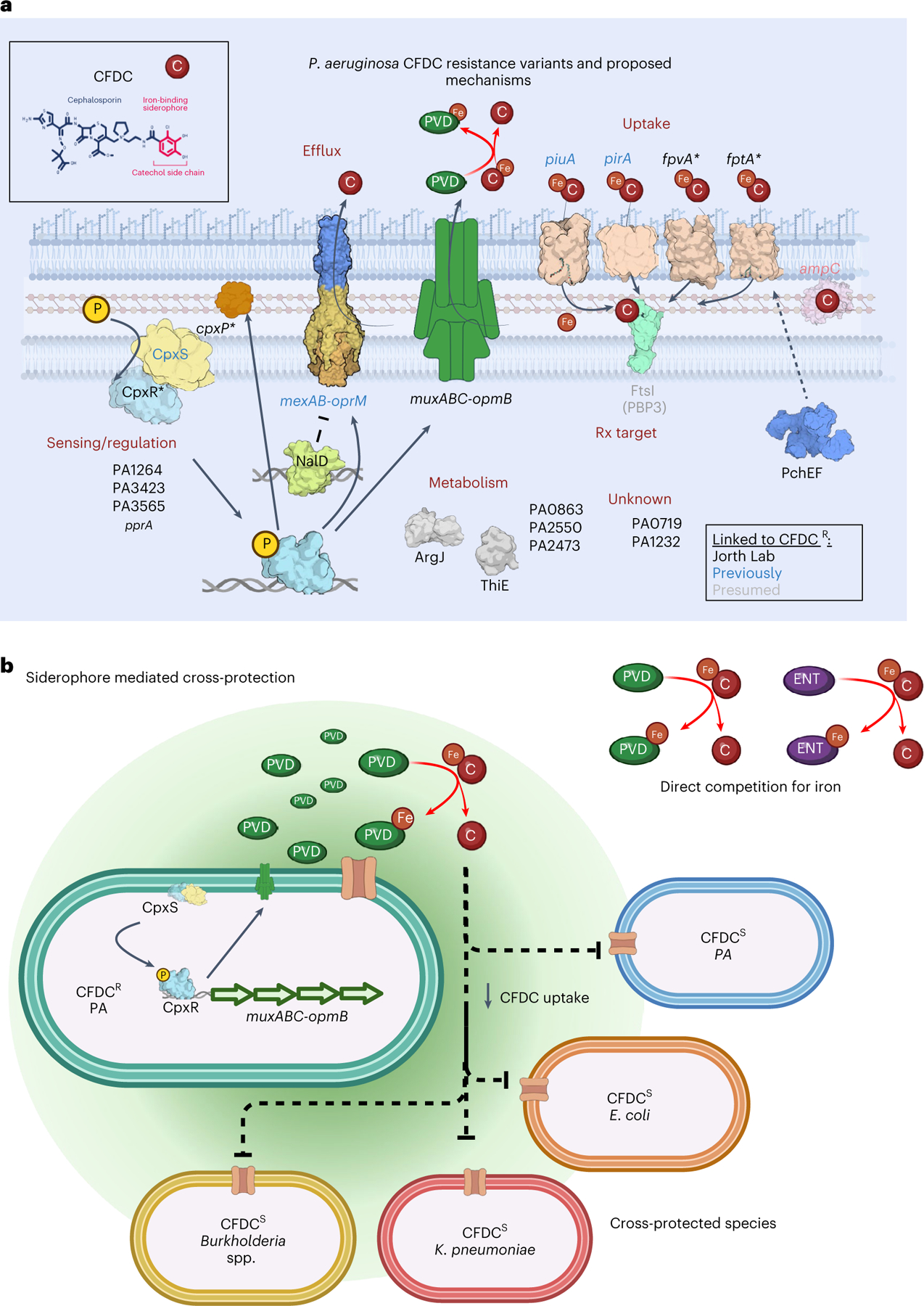
The antibiotic cefiderocol hijacks iron transporters to facilitate its uptake and resists β-lactamase degradation. While effective, resistance has been detected clinically with unknown mechanisms. Here, using experimental evolution, we identified cefiderocol resistance mutations in Pseudomonas aeruginosa. Resistance was multifactorial in host-mimicking growth media, led to multidrug resistance and paid fitness costs in cefiderocol-free environments. However, kin selection drove some resistant populations to cross-protect susceptible individuals from killing by increasing pyoverdine secretion via a two-component sensor mutation. While pyochelin sensitized P. aeruginosa to cefiderocol killing, pyoverdine and the enterobacteria siderophore enterobactin displaced iron from cefiderocol, preventing uptake by susceptible cells. Among 113 P. aeruginosa intensive care unit clinical isolates, pyoverdine production directly correlated with cefiderocol tolerance, and high pyoverdine producing isolates cross-protected susceptible P. aeruginosa and other Gram-negative bacteria. These in vitro data show that antibiotic cross-protection can occur via degradation-independent mechanisms and siderophores can serve unexpected protective cooperative roles in polymicrobial communities.
© 2024. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures







Update of
-
Evolved bacterial siderophore-mediated antibiotic cross-protection.Res Sq [Preprint]. 2023 May 18:rs.3.rs-2644953. doi: 10.21203/rs.3.rs-2644953/v1. Res Sq. 2023. Update in: Nat Microbiol. 2024 Mar;9(3):631-646. doi: 10.1038/s41564-024-01601-4. PMID: 37292841 Free PMC article. Updated. Preprint.
References
-
- Tacconelli E et al. Surveillance for control of antimicrobial resistance. Lancet Infect. Dis 18, e99–e106 (2018). - PubMed
-
- Mittal R, Aggarwal S, Sharma S, Chhibber S & Harjai K Urinary tract infections caused by Pseudomonas aeruginosa: a minireview. J. Infect. Public Health 2, 101–111 (2009). - PubMed
MeSH terms
Substances
Grants and funding
- JORTH19P0/Cystic Fibrosis Foundation (CF Foundation)
- K22 AI127473/AI/NIAID NIH HHS/United States
- R01 AI146425/AI/NIAID NIH HHS/United States
- R01Hl136143/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R01 HL136143/HL/NHLBI NIH HHS/United States
- R01AI14642/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R21 AI151362/AI/NIAID NIH HHS/United States
- JORTH17F5/Cystic Fibrosis Foundation (CF Foundation)
- MILESI21F0/Cystic Fibrosis Foundation (CF Foundation)
- R21AI151362/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- K22AI127473/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
