

This is a preprint.
Cover Crop Root Exudates Impact Soil Microbiome Functional Trajectories in Agricultural Soils
- PMID: 38410449
- PMCID: PMC10896397
- DOI: 10.21203/rs.3.rs-3956430/v1
Cover Crop Root Exudates Impact Soil Microbiome Functional Trajectories in Agricultural Soils
Update in
-
Cover crop root exudates impact soil microbiome functional trajectories in agricultural soils.Microbiome. 2024 Sep 28;12(1):183. doi: 10.1186/s40168-024-01886-x. Microbiome. 2024. PMID: 39342284 Free PMC article.
Abstract
Background: Cover cropping is an agricultural practice that uses secondary crops to support the growth of primary crops through various mechanisms including erosion control, weed suppression, nutrient management, and enhanced biodiversity. Cover crops may elicit some of these ecosystem services through chemical interactions with the soil microbiome via root exudation, or the release of plant metabolites from roots. Phytohormones are one metabolite type exuded by plants that activate the rhizosphere microbiome, yet managing this chemical interaction remains an untapped mechanism for optimizing plant-soil microbiome interactions. Currently, there is limited understanding on the diversity of cover crop phytohormone root exudation patterns and how these chemical messages selectively enrich specific microbial taxa and functionalities in agricultural soils.
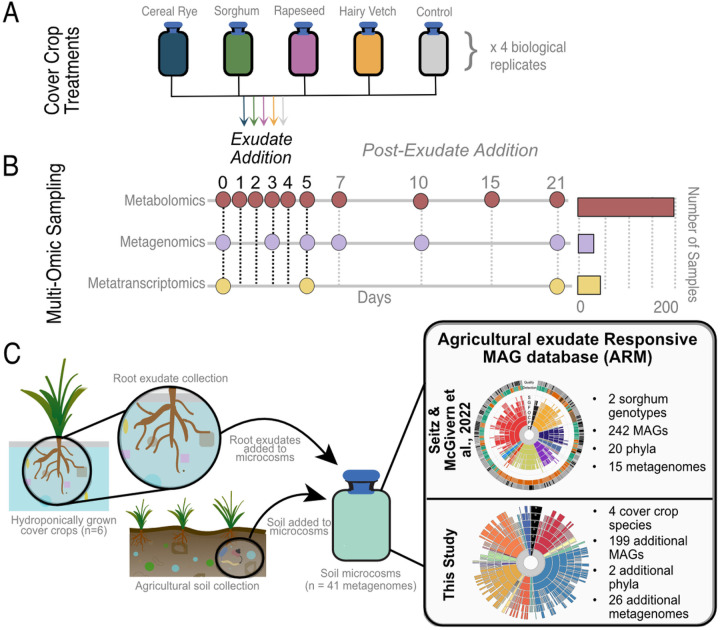
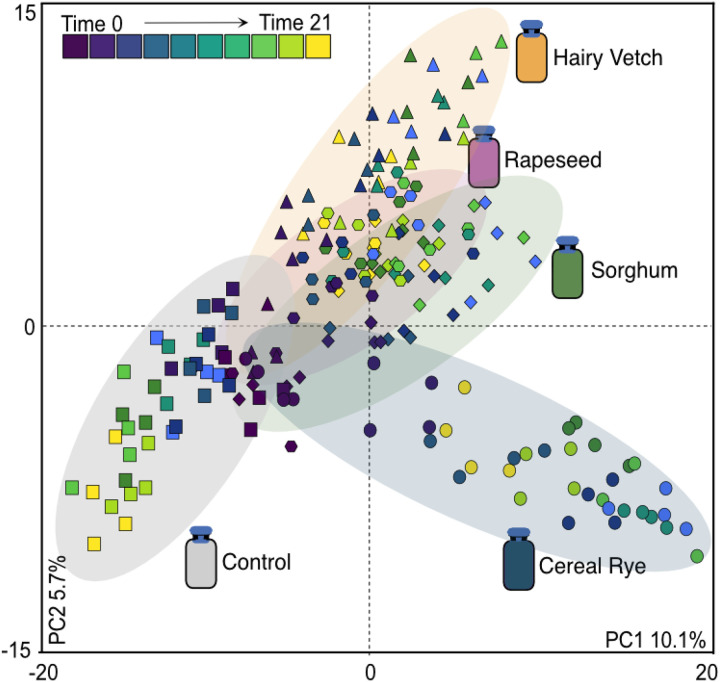
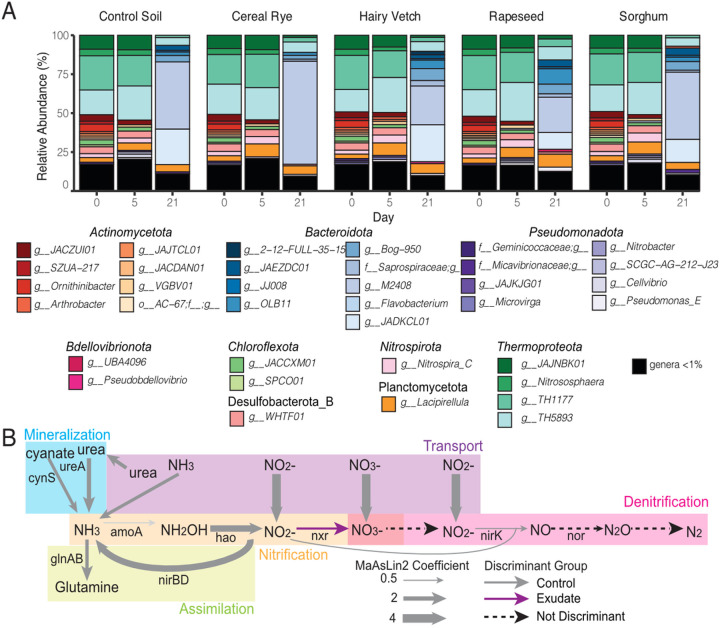
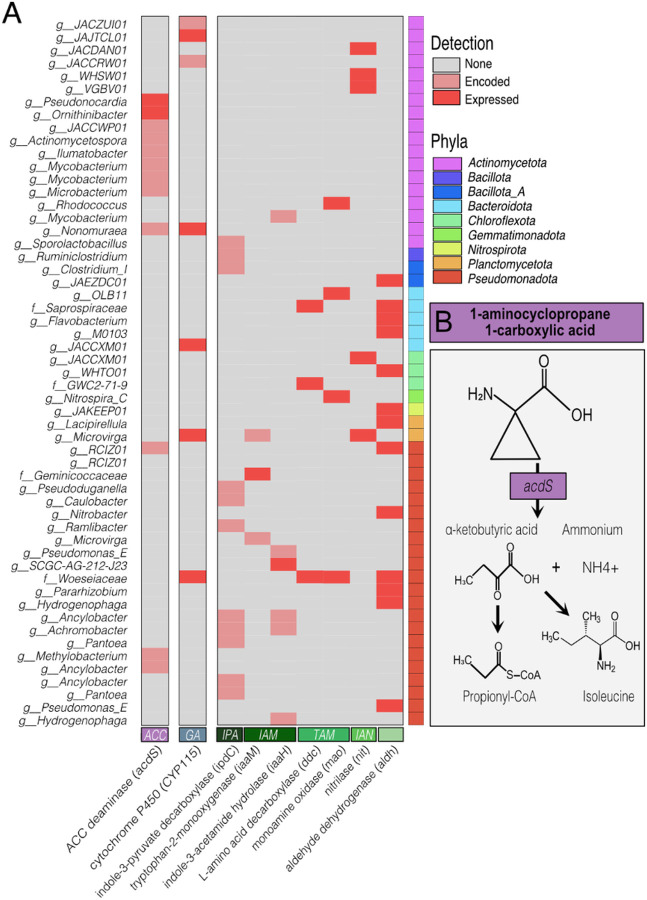
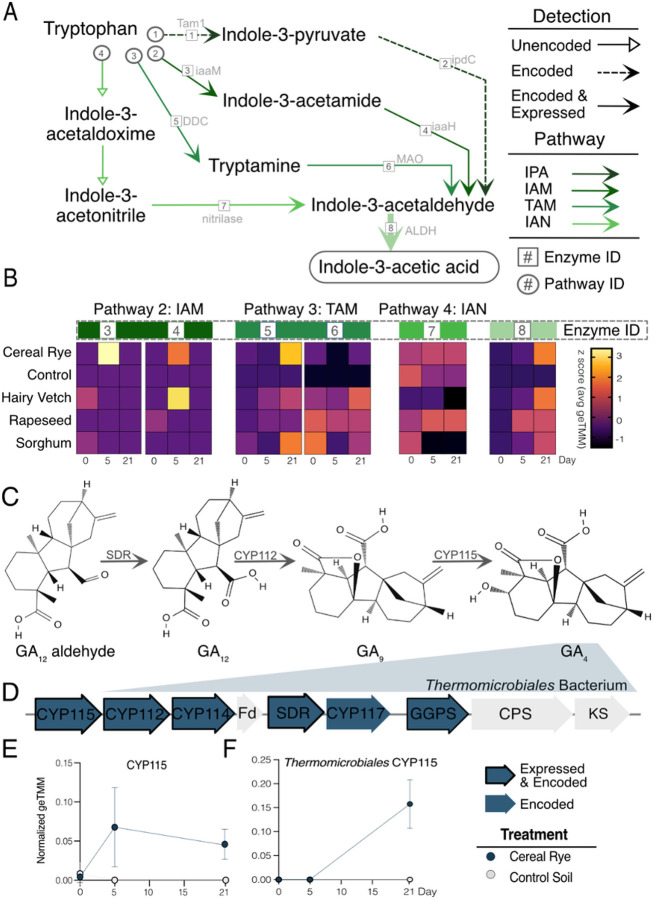
Results: Here, we link variability in cover crop root exudate composition to changes in soil microbiome functionality. Exudate chemical profiles from 4 cover crop species (Sorghum bicolor, Vicia villosa, Brassica napus, and Secale cereal) were used as the chemical inputs to decipher microbial responses. These distinct exudate profiles, along with a no exudate control, were amended to agricultural soil microcosms with microbial responses tracked over time using metabolomes and genome-resolved metatranscriptomes. Our findings illustrated microbial metabolic patterns were unique in response to cover crop exudate inputs over time, particularly by sorghum and cereal rye amended microcosms where we identify novel microbial members (at the genera and family level) who produced IAA and GA4 over time. We also identify broad changes in microbial nitrogen cycling in response chemical inputs.
Conclusions: We highlight that root exudate amendments alter microbial community function and phytohormone metabolisms, particularly in response to root exudates isolated from cereal rye and sorghum plants. Additionally, we constructed a soil microbial genomic catalog of microorganisms responding to commonly used cover crops, a public resource for agriculturally-relevant microbes. Many of our exudate-stimulated microorganisms are representatives from poorly characterized or novel taxa, highlighting the yet to be discovered metabolic reservoir harbored in agricultural soils. Our findings emphasize the tractability of high-resolution multiomics approaches to investigate processes relevant for agricultural soils, opening the possibility of targeting specific soil biogeochemical outcomes through biological precision agricultural practices that use cover crops and the microbiome as levers for enhanced crop production.
Keywords: Brassica napus; Phytohormone; Secale cereal; Sorghum bicolor; Vicia villosa; liquid chromatography mass spectrometry (LC-MS); metagenome assembled genome (MAG); metatranscriptomics; plant growth promoting.
Conflict of interest statement
Competing interests: The authors have no competing interests. Additional Declarations: No competing interests reported.
Figures


References
-
- SARE: Annual Report 2019–2020 National Cover Crop Survey In. Sustainable Agriculture Research and Education (SARE); 2020.
-
- Mitchell JP, Shrestha A, Mathesius K, Scow KM, Southard RJ, Haney RL, et al. Cover cropping and no-tillage improve soil health in an arid irrigated cropping system in California’s San Joaquin Valley, USA. Soil and Tillage Research. 2017;165:325–35; doi: 10.1016/j.still.2016.09.001. - DOI
-
- Vukicevich E, Lowery T, Bowen P Úrbez-Torres JR, Hart M. Cover crops to increase soil microbial diversity and mitigate decline in perennial agriculture. A review. Agronomy for Sustainable Development. 2016;36(3):48; doi: 10.1007/s13593-016-0385-7. - DOI
-
- Strickland MS, Thomason WE, Avera B, Franklin J, Minick K, Yamada S, et al. Short-Term Effects of Cover Crops on Soil Microbial Characteristics and Biogeochemical Processes across Actively Managed Farms. Agrosystems, Geosciences & Environment. 2019;2(1):180064; doi: 10.2134/age2018.12.0064. - DOI
-
- Shackelford GE K Rodd; Dicks Lynn V. Effects of cover crops on multiple ecosystem services: Ten meta-analyses of data from arable farmland in California and the Mediterranean. Land Use Policy. 2019;88.
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
