

The Caenorhabditis elegans proteome response to two protective Pseudomonas symbionts
- PMID: 38411078
- PMCID: PMC11005407
- DOI: 10.1128/mbio.03463-23
The Caenorhabditis elegans proteome response to two protective Pseudomonas symbionts
Abstract
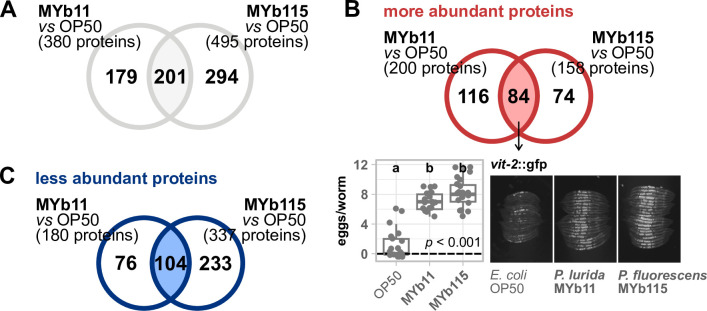
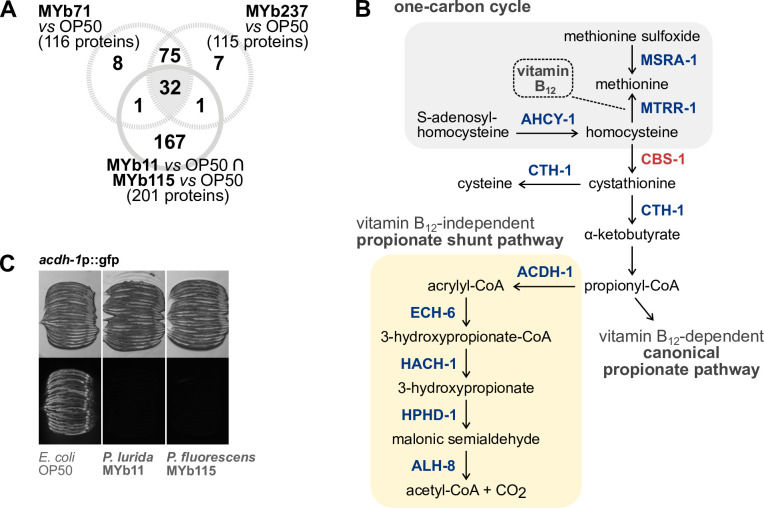
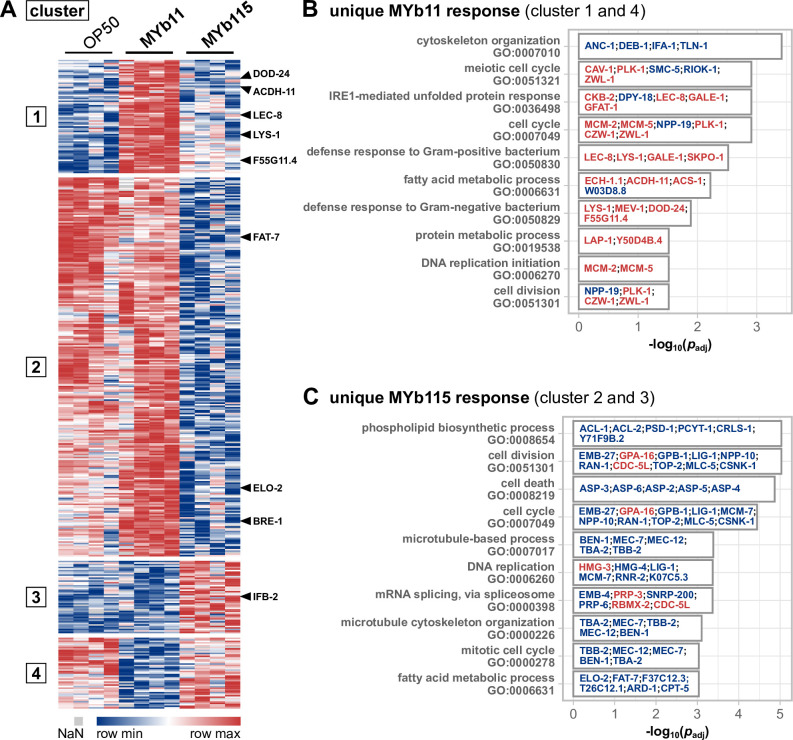
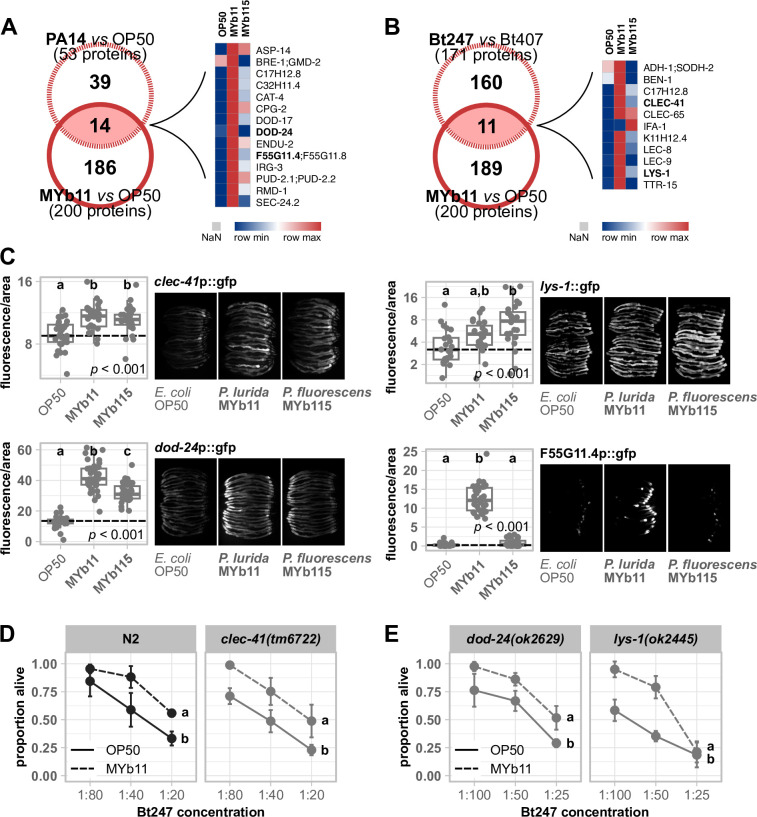
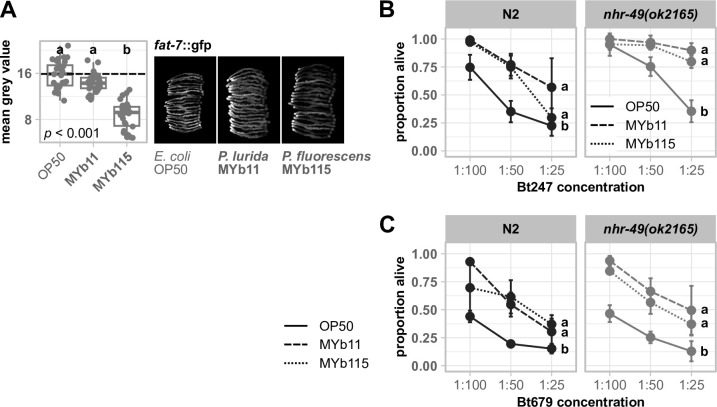
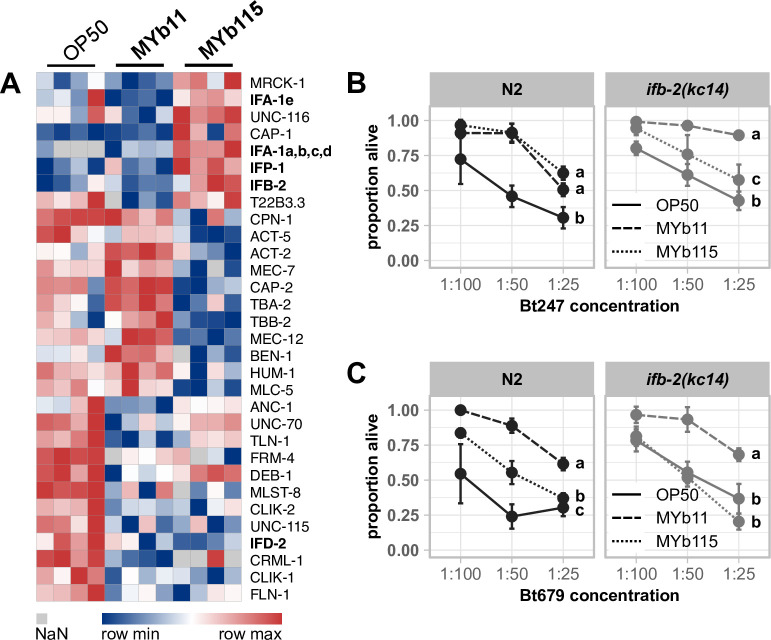
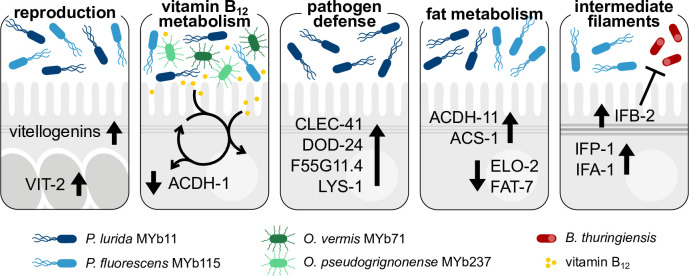
The Caenorhabditis elegans natural microbiota isolates Pseudomonas lurida MYb11 and Pseudomonas fluorescens MYb115 protect the host against pathogens through distinct mechanisms. While P. lurida produces an antimicrobial compound and directly inhibits pathogen growth, P. fluorescens MYb115 protects the host without affecting pathogen growth. It is unknown how these two protective microbes affect host biological processes. We used a proteomics approach to elucidate the C. elegans response to MYb11 and MYb115. We found that both Pseudomonas isolates increase vitellogenin protein production in young adults, which confirms previous findings on the effect of microbiota on C. elegans reproductive timing. Moreover, the C. elegans responses to MYb11 and MYb115 exhibit common signatures with the response to other vitamin B12-producing bacteria, emphasizing the importance of vitamin B12 in C. elegans-microbe metabolic interactions. We further analyzed signatures in the C. elegans response specific to MYb11 or MYb115. We provide evidence for distinct modifications in lipid metabolism by both symbiotic microbes. We could identify the activation of host-pathogen defense responses as an MYb11-specific proteome signature and provide evidence that the intermediate filament protein IFB-2 is required for MYb115-mediated protection. These results indicate that MYb11 not only produces an antimicrobial compound but also activates host antimicrobial defenses, which together might increase resistance to infection. In contrast, MYb115 affects host processes such as lipid metabolism and cytoskeleton dynamics, which might increase host tolerance to infection. Overall, this study pinpoints proteins of interest that form the basis for additional exploration into the mechanisms underlying C. elegans microbiota-mediated protection from pathogen infection and other microbiota-mediated traits.IMPORTANCESymbiotic bacteria can defend their host against pathogen infection. While some protective symbionts directly interact with pathogenic bacteria, other protective symbionts elicit a response in the host that improves its own pathogen defenses. To better understand how a host responds to protective symbionts, we examined which host proteins are affected by two protective Pseudomonas bacteria in the model nematode Caenorhabditis elegans. We found that the C. elegans response to its protective symbionts is manifold, which was reflected in changes in proteins that are involved in metabolism, the immune system, and cell structure. This study provides a foundation for exploring the contribution of the host response to symbiont-mediated protection from pathogen infection.
Keywords: Caenorhabditis elegans; Pseudomonas; microbiota; microbiota-mediated protection; proteome.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Dirksen P, Marsh SA, Braker I, Heitland N, Wagner S, Nakad R, Mader S, Petersen C, Kowallik V, Rosenstiel P, Félix M-A, Schulenburg H. 2016. The native microbiome of the nematode Caenorhabditis elegans: gateway to a new host-microbiome model. BMC Biol 14:38. doi:10.1186/s12915-016-0258-1 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
