

Ribosomal quality control factors inhibit repeat-associated non-AUG translation from GC-rich repeats
- PMID: 38412259
- PMCID: PMC11162809
- DOI: 10.1093/nar/gkae137
Ribosomal quality control factors inhibit repeat-associated non-AUG translation from GC-rich repeats
Abstract
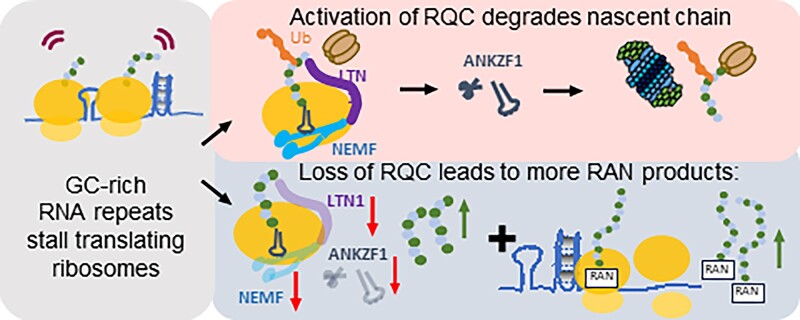
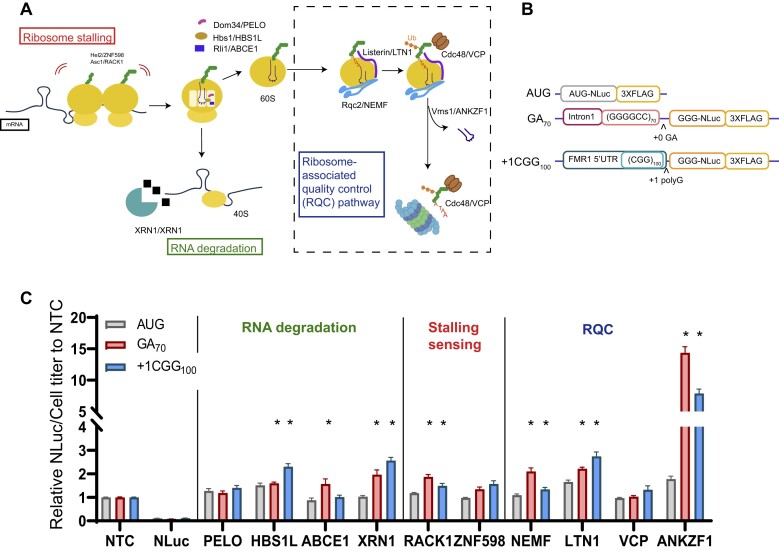
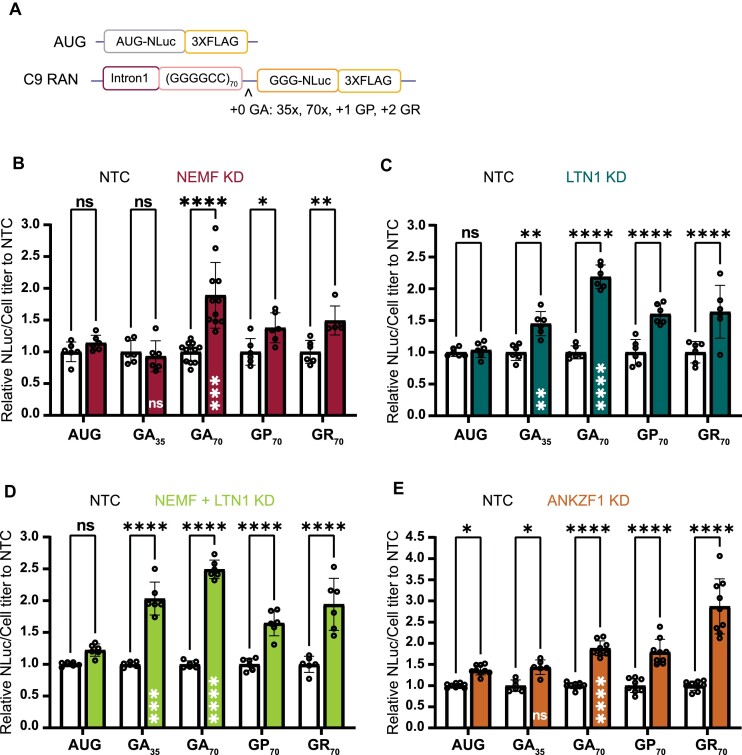
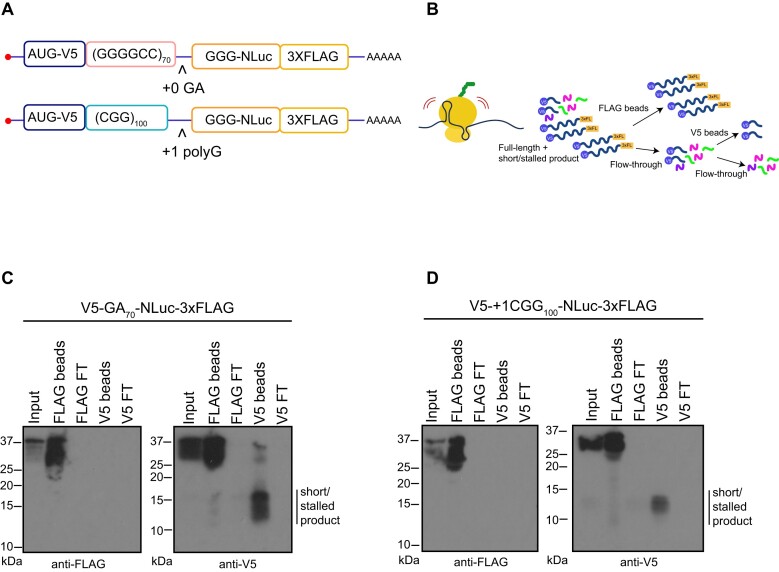
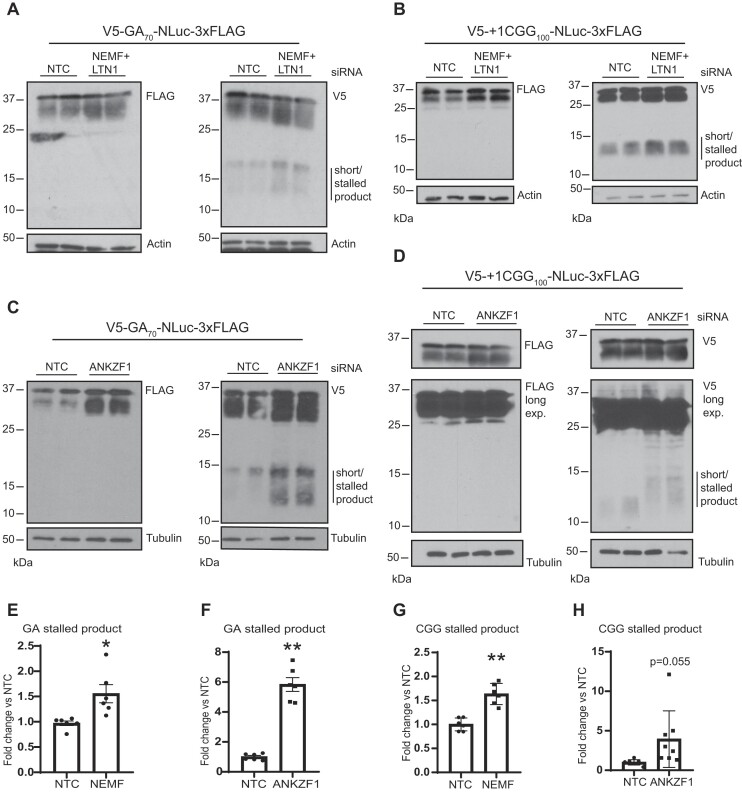
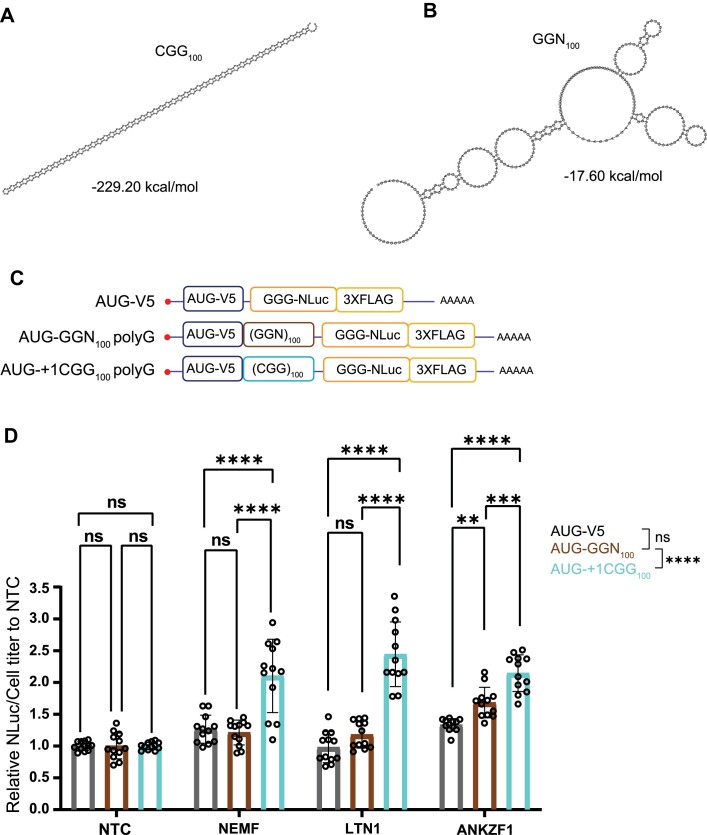
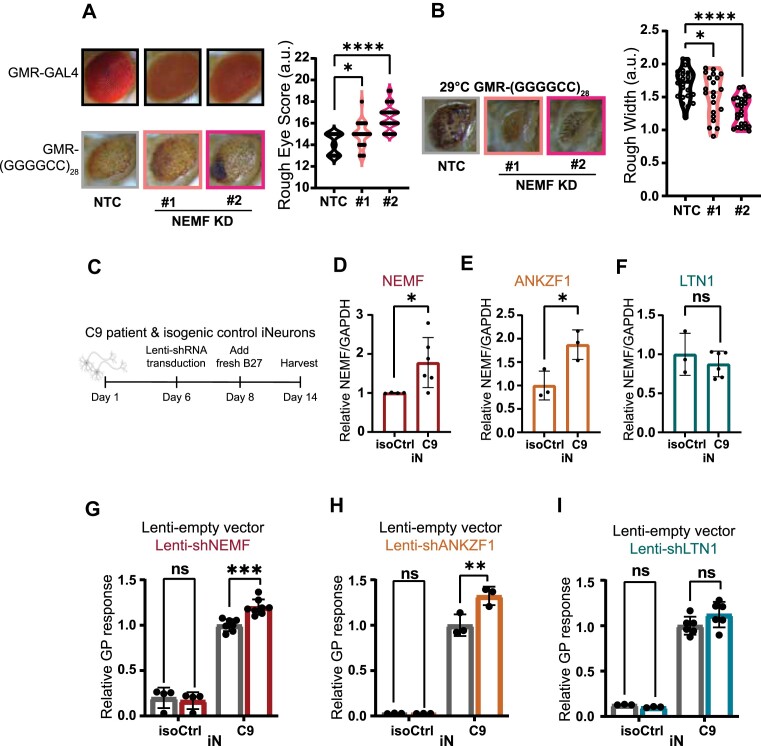
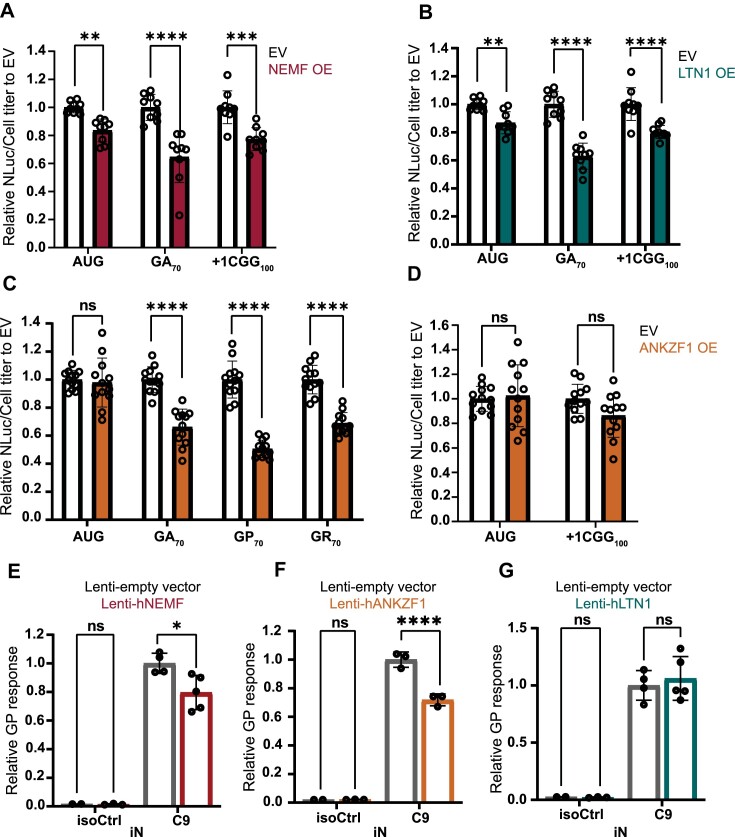
A GGGGCC (G4C2) hexanucleotide repeat expansion in C9ORF72 causes amyotrophic lateral sclerosis and frontotemporal dementia (C9ALS/FTD), while a CGG trinucleotide repeat expansion in FMR1 leads to the neurodegenerative disorder Fragile X-associated tremor/ataxia syndrome (FXTAS). These GC-rich repeats form RNA secondary structures that support repeat-associated non-AUG (RAN) translation of toxic proteins that contribute to disease pathogenesis. Here we assessed whether these same repeats might trigger stalling and interfere with translational elongation. We find that depletion of ribosome-associated quality control (RQC) factors NEMF, LTN1 and ANKZF1 markedly boost RAN translation product accumulation from both G4C2 and CGG repeats while overexpression of these factors reduces RAN production in both reporter assays and C9ALS/FTD patient iPSC-derived neurons. We also detected partially made products from both G4C2 and CGG repeats whose abundance increased with RQC factor depletion. Repeat RNA sequence, rather than amino acid content, is central to the impact of RQC factor depletion on RAN translation-suggesting a role for RNA secondary structure in these processes. Together, these findings suggest that ribosomal stalling and RQC pathway activation during RAN translation inhibits the generation of toxic RAN products. We propose augmenting RQC activity as a therapeutic strategy in GC-rich repeat expansion disorders.
© The Author(s) 2024. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Update of
-
Ribosomal quality control factors inhibit repeat-associated non-AUG translation from GC-rich repeats.bioRxiv [Preprint]. 2023 Jun 7:2023.06.07.544135. doi: 10.1101/2023.06.07.544135. bioRxiv. 2023. Update in: Nucleic Acids Res. 2024 Jun 10;52(10):5928-5949. doi: 10.1093/nar/gkae137. PMID: 37333274 Free PMC article. Updated. Preprint.
References
-
- Koob M.D., Moseley M.L., Schut L.J., Benzow K.A., Bird T.D., Day J.W., Ranum L.P.W. An untranslated CTG expansion causes a novel form of spinocerebellar ataxia (SCA8). Nat. Genet. 1999; 21:379–384. - PubMed
-
- Lander E.S., Linton L.M., Birren B., Nusbaum C., Zody M.C., Baldwin J., Devon K., Dewar K., Doyle M., Fitzhugh W. et al. . Initial sequencing and analysis of the human genome: international Human Genome Sequencing Consortium. Nature. 2001; 409:860–921. - PubMed
-
- Liquori C.L., Ricker K., Moseley M.L., Jacobsen J.F., Kress W., Naylor S.L., Day J.W., Ranum L.P.W. Myotonic dystrophy type 2 caused by a CCTG expansion in intron I of ZNF9. Science. 2001; 293:864–867. - PubMed
-
- Verkerk A.J.M.H., Pieretti M., Sutcliffe J.S., Fu Y.H., Kuhl D.P.A., Pizzuti A., Reiner O., Richards S., Victoria M.F., Zhang F. et al. . Identification of a gene (FMR-1) containing a CGG repeat coincident with a breakpoint cluster region exhibiting length variation in fragile X syndrome. Cell. 1991; 65:905–914. - PubMed
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
