

TopBP1 utilises a bipartite GINS binding mode to support genome replication
- PMID: 38413589
- PMCID: PMC10899662
- DOI: 10.1038/s41467-024-45946-0
TopBP1 utilises a bipartite GINS binding mode to support genome replication
Abstract
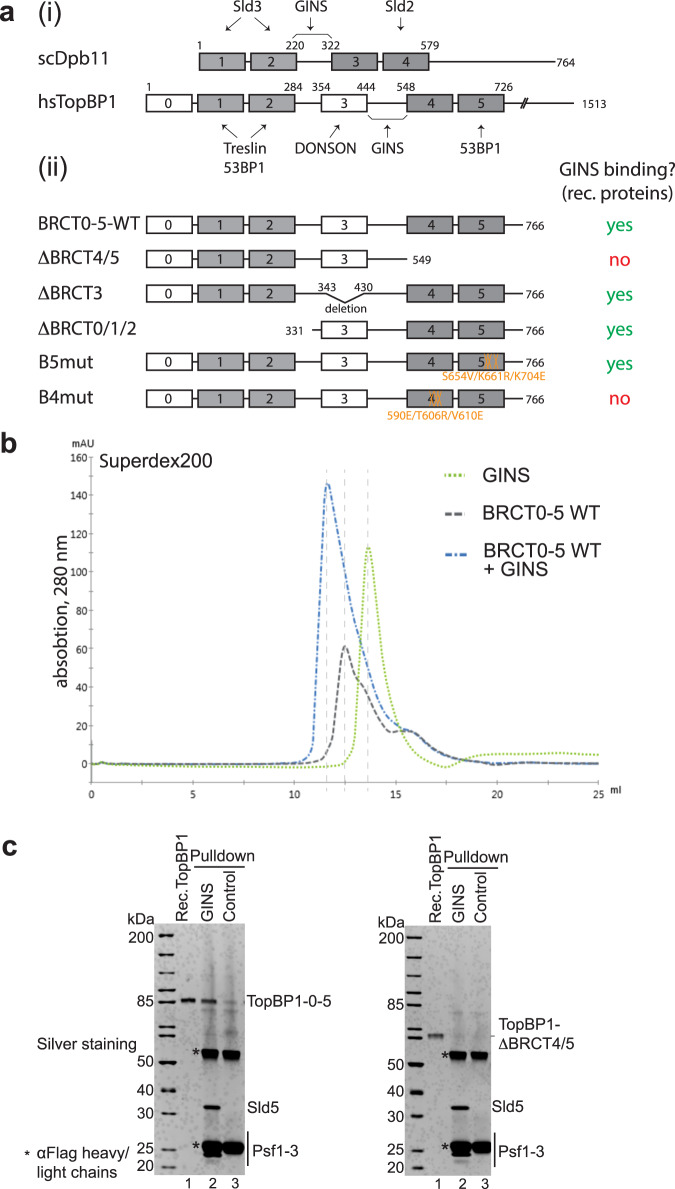
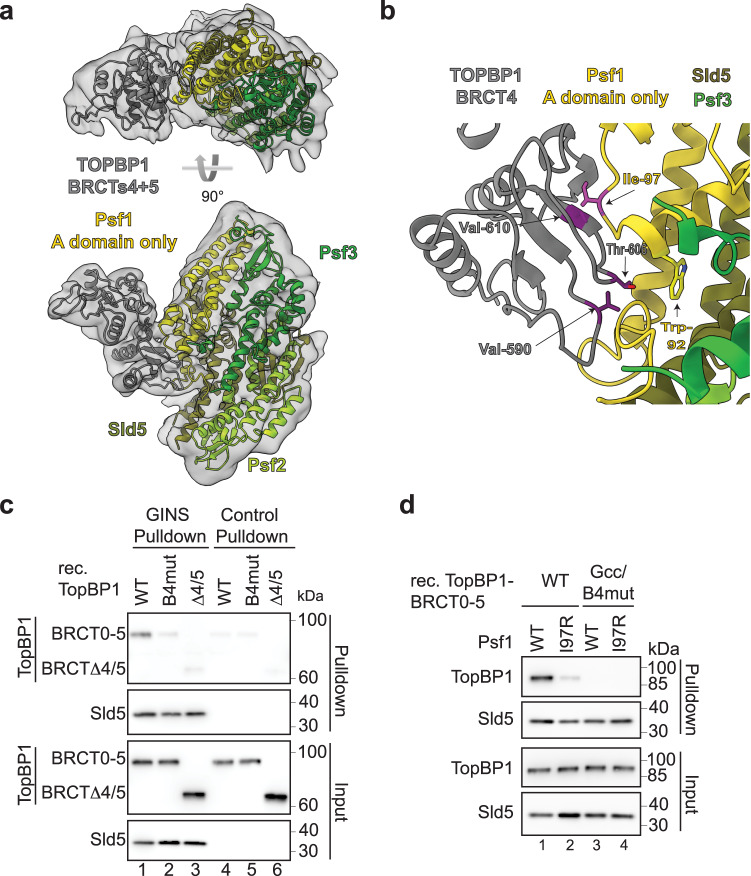
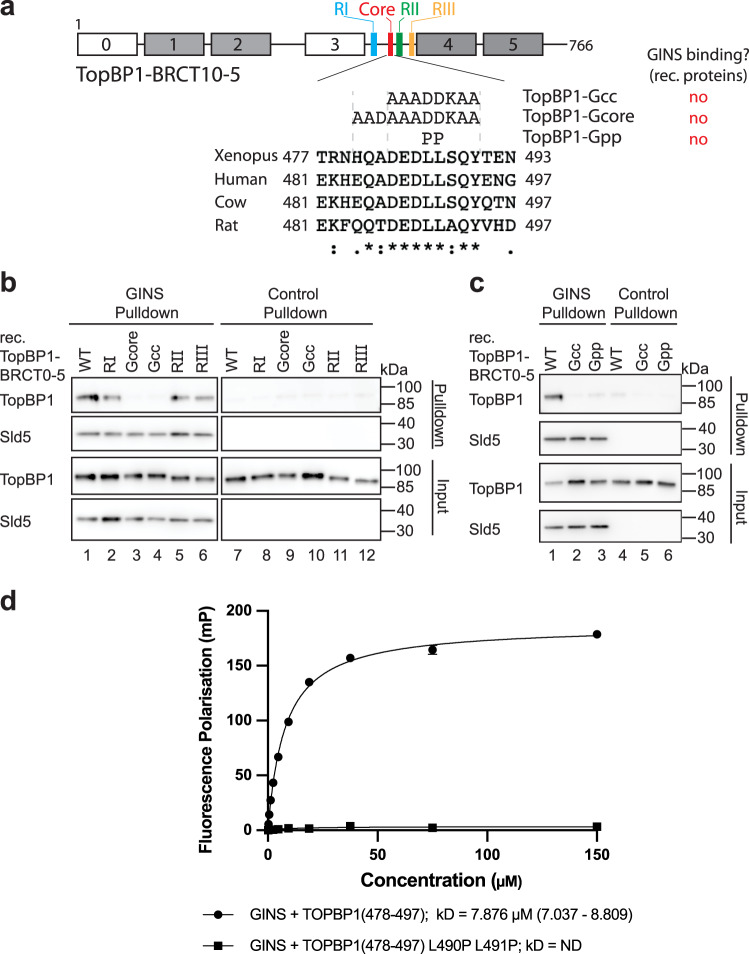
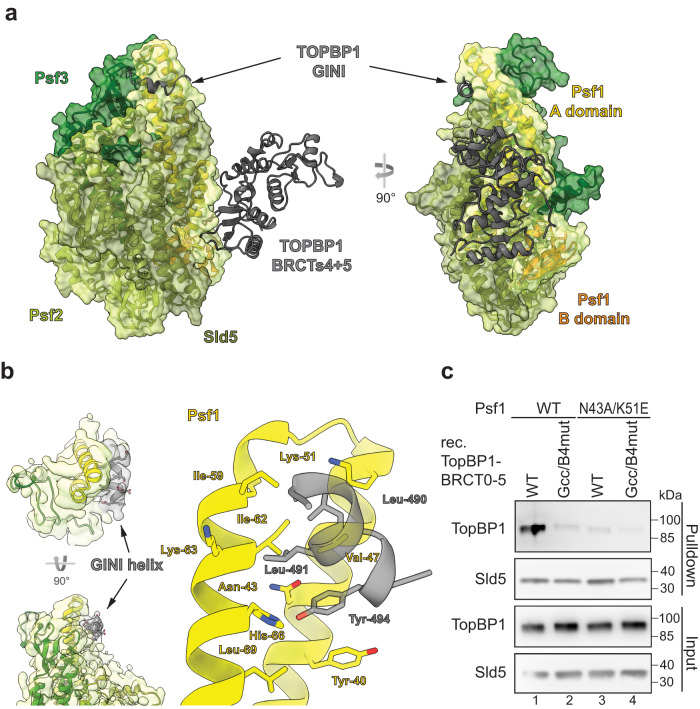
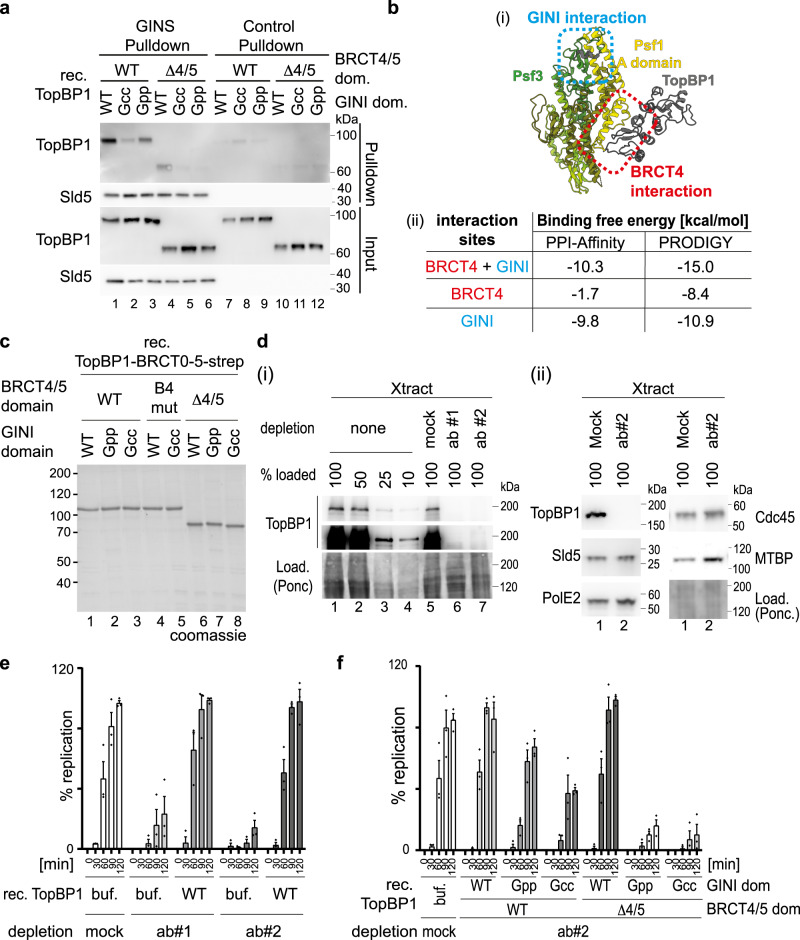
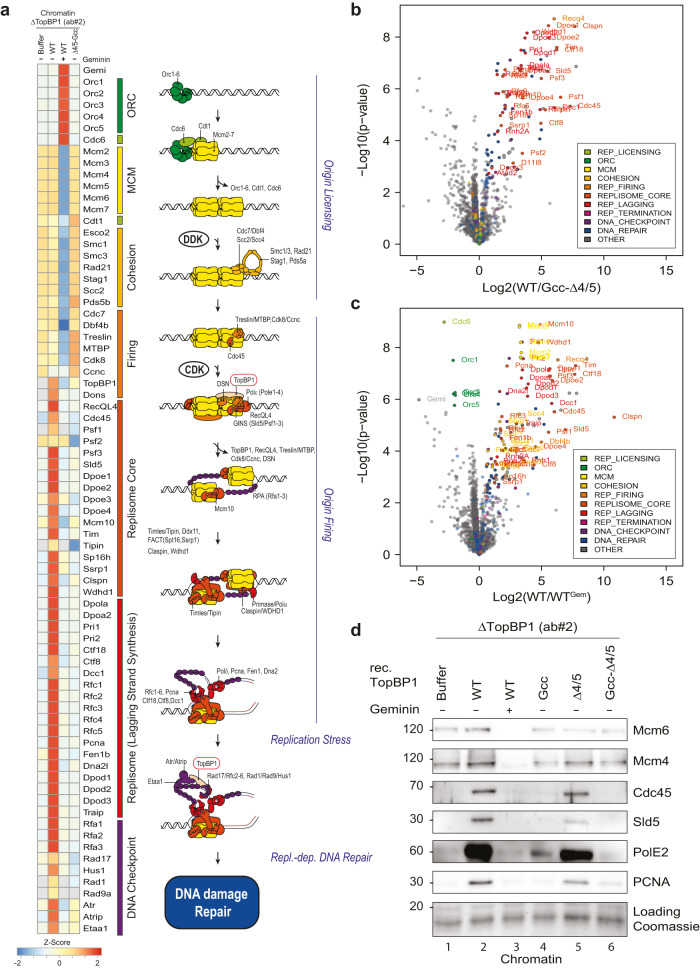
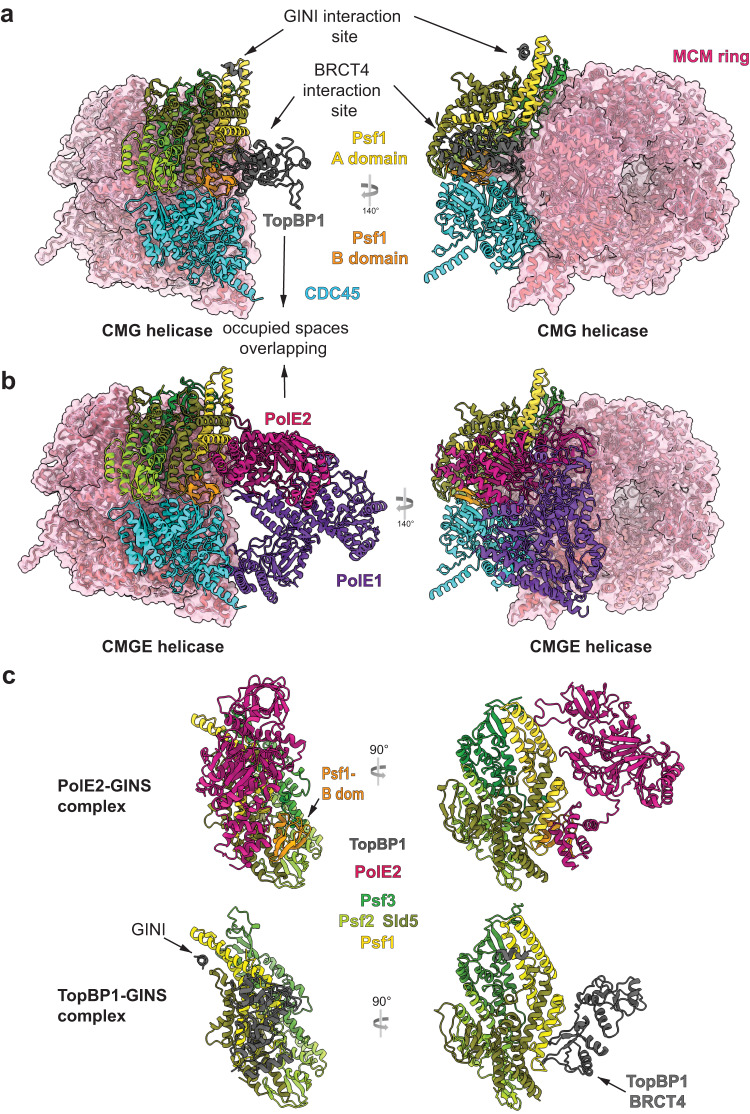
Activation of the replicative Mcm2-7 helicase by loading GINS and Cdc45 is crucial for replication origin firing, and as such for faithful genetic inheritance. Our biochemical and structural studies demonstrate that the helicase activator GINS interacts with TopBP1 through two separate binding surfaces, the first involving a stretch of highly conserved amino acids in the TopBP1-GINI region, the second a surface on TopBP1-BRCT4. The two surfaces bind to opposite ends of the A domain of the GINS subunit Psf1. Mutation analysis reveals that either surface is individually able to support TopBP1-GINS interaction, albeit with reduced affinity. Consistently, either surface is sufficient for replication origin firing in Xenopus egg extracts and becomes essential in the absence of the other. The TopBP1-GINS interaction appears sterically incompatible with simultaneous binding of DNA polymerase epsilon (Polε) to GINS when bound to Mcm2-7-Cdc45, although TopBP1-BRCT4 and the Polε subunit PolE2 show only partial competitivity in binding to Psf1. Our TopBP1-GINS model improves the understanding of the recently characterised metazoan pre-loading complex. It further predicts the coordination of three molecular origin firing processes, DNA polymerase epsilon arrival, TopBP1 ejection and GINS integration into Mcm2-7-Cdc45.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Li N, et al. Structure of the eukaryotic MCM complex at 3.8 A. Nature. 2015;524:186–191. - PubMed
MeSH terms
Substances
Grants and funding
- SFB1430; 424228829/Deutsche Forschungsgemeinschaft (German Research Foundation)
- RTG1739/Deutsche Forschungsgemeinschaft (German Research Foundation)
- FOR2800/Deutsche Forschungsgemeinschaft (German Research Foundation)
- SFB1430; 424228829/Deutsche Forschungsgemeinschaft (German Research Foundation)
- INST 20876/322-1 FUGG/Deutsche Forschungsgemeinschaft (German Research Foundation)
- SFB1430; 424228829/Deutsche Forschungsgemeinschaft (German Research Foundation)
- EXC-390677874-RESOLV/Deutsche Forschungsgemeinschaft (German Research Foundation)
- INST - 436586093/Deutsche Forschungsgemeinschaft (German Research Foundation)
- INST 20876/322-1 FUGG/Deutsche Forschungsgemeinschaft (German Research Foundation)
- SFB1430; 424228829/Deutsche Forschungsgemeinschaft (German Research Foundation)
- C302/A14532/Cancer Research UK (CRUK)
- C302/A24386/Cancer Research UK (CRUK)
- C302/A14532/Cancer Research UK (CRUK)
- C302/A24386/Cancer Research UK (CRUK)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
