

Parkinson's disease and schizophrenia interactomes contain temporally distinct gene clusters underlying comorbid mechanisms and unique disease processes
- PMID: 38413605
- PMCID: PMC10899210
- DOI: 10.1038/s41537-024-00439-3
Parkinson's disease and schizophrenia interactomes contain temporally distinct gene clusters underlying comorbid mechanisms and unique disease processes
Erratum in
-
Author Correction: Parkinson's disease and schizophrenia interactomes contain temporally distinct gene clusters underlying comorbid mechanisms and unique disease processes.Schizophrenia (Heidelb). 2024 Mar 13;10(1):33. doi: 10.1038/s41537-024-00455-3. Schizophrenia (Heidelb). 2024. PMID: 38480791 Free PMC article. No abstract available.
Abstract
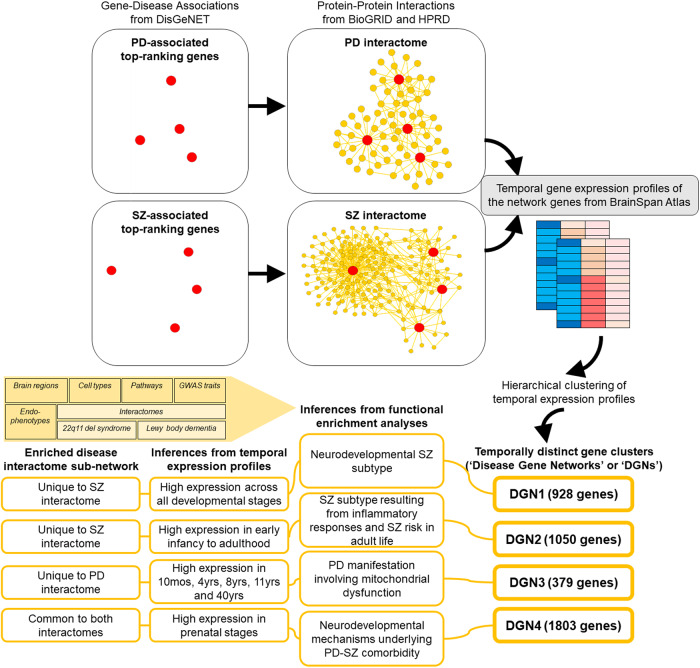
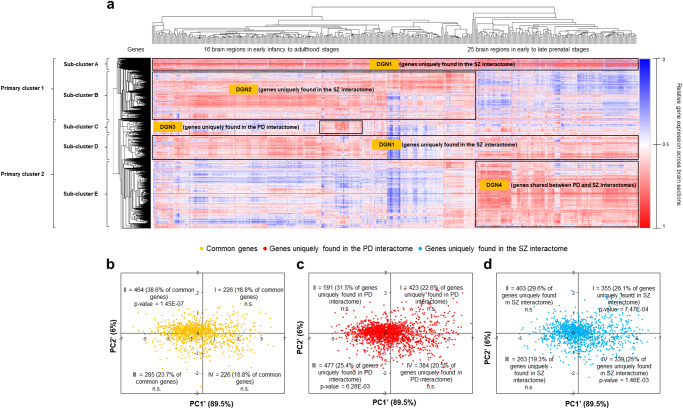
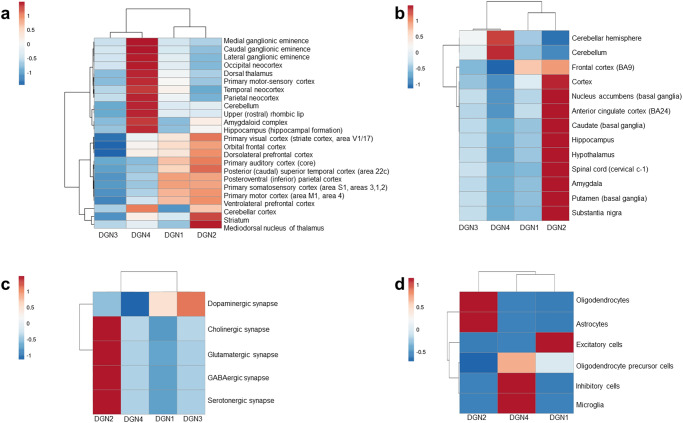
Genome-wide association studies suggest significant overlaps in Parkinson's disease (PD) and schizophrenia (SZ) risks, but the underlying mechanisms remain elusive. The protein-protein interaction network ('interactome') plays a crucial role in PD and SZ and can incorporate their spatiotemporal specificities. Therefore, to study the linked biology of PD and SZ, we compiled PD- and SZ-associated genes from the DisGeNET database, and constructed their interactomes using BioGRID and HPRD. We examined the interactomes using clustering and enrichment analyses, in conjunction with the transcriptomic data of 26 brain regions spanning foetal stages to adulthood available in the BrainSpan Atlas. PD and SZ interactomes formed four gene clusters with distinct temporal identities (Disease Gene Networks or 'DGNs'1-4). DGN1 had unique SZ interactome genes highly expressed across developmental stages, corresponding to a neurodevelopmental SZ subtype. DGN2, containing unique SZ interactome genes expressed from early infancy to adulthood, correlated with an inflammation-driven SZ subtype and adult SZ risk. DGN3 contained unique PD interactome genes expressed in late infancy, early and late childhood, and adulthood, and involved in mitochondrial pathways. DGN4, containing prenatally-expressed genes common to both the interactomes, involved in stem cell pluripotency and overlapping with the interactome of 22q11 deletion syndrome (comorbid psychosis and Parkinsonism), potentially regulates neurodevelopmental mechanisms in PD-SZ comorbidity. Our findings suggest that disrupted neurodevelopment (regulated by DGN4) could expose risk windows in PD and SZ, later elevating disease risk through inflammation (DGN2). Alternatively, variant clustering in DGNs may produce disease subtypes, e.g., PD-SZ comorbidity with DGN4, and early/late-onset SZ with DGN1/DGN2.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Spatial and functional profiles distinguish target sets of Parkinson's disease and antipsychotic drugs with different clinical effects.Transl Psychiatry. 2025 Apr 4;15(1):124. doi: 10.1038/s41398-025-03351-1. Transl Psychiatry. 2025. PMID: 40185727 Free PMC article.
-
A mouse model of 22q11.2 deletions: Molecular and behavioral signatures of Parkinson's disease and schizophrenia.Sci Adv. 2018 Aug 15;4(8):eaar6637. doi: 10.1126/sciadv.aar6637. eCollection 2018 Aug. Sci Adv. 2018. PMID: 30116778 Free PMC article.
-
Author Correction: Parkinson's disease and schizophrenia interactomes contain temporally distinct gene clusters underlying comorbid mechanisms and unique disease processes.Schizophrenia (Heidelb). 2024 Mar 13;10(1):33. doi: 10.1038/s41537-024-00455-3. Schizophrenia (Heidelb). 2024. PMID: 38480791 Free PMC article. No abstract available.
-
Neurodevelopmental disturbances in schizophrenia: evidence from genetic and environmental factors.J Neural Transm (Vienna). 2023 Mar;130(3):195-205. doi: 10.1007/s00702-022-02567-5. Epub 2022 Nov 12. J Neural Transm (Vienna). 2023. PMID: 36370183 Free PMC article. Review.
-
Toward the identification of peripheral epigenetic biomarkers of schizophrenia.J Neurogenet. 2014 Mar-Jun;28(1-2):41-52. doi: 10.3109/01677063.2014.892485. Epub 2014 Apr 22. J Neurogenet. 2014. PMID: 24702539 Free PMC article. Review.
References
-
- Morris, S., Olichney, J. & Corey-Bloom, J. In Seminars in clinical neuropsychiatry. 51–60. - PubMed
LinkOut - more resources
Full Text Sources
